PDF version: global highlights in neuroengineering 2005-2018 – logan thrasher collins
Optogenetic stimulation using ChR2
(Boyden, Zhang, Bamberg, Nagel, & Deisseroth, 2005)
- Ed Boyden, Karl Deisseroth, and colleagues developed optogenetics, a revolutionary technique for stimulating neural activity.
- Optogenetics involves engineering neurons to express light-gated ion channels. The first channel used for this purpose was ChR2 (a protein originally found in bacteria which responds to blue light). In this way, a neuron exposed to an appropriate wavelength of light will be stimulated.
- Over time, optogenetics has gained a place as an essential experimental tool for neuroscientists across the world. It has been expanded upon and improved in numerous ways and has even allowed control of animal behavior via implanted fiber optics and other light sources. Optogenetics may eventually be used in the development of improved brain-computer interfaces.

Blue Brain Project cortical column simulation
(Markram, 2006)
- In the early stages of the Blue Brain Project, neuronal cell types from the layers of the rat neocortex were reconstructed. Furthermore, their electrophysiology was experimentally characterized.
- Next, a virtual neocortical column with about 10,000 multicompartmental Hodgkin-Huxley-type neurons and over ten million synapses was built. Its connectivity was defined according the patterns of connectivity found in biological rats, (though this involved the numbers of inputs and outputs quantified for given cell types rather than explicit wiring). In addition, the spatial distributions of boutons forming synaptic terminals upon target cells reflected biological data.
- The cortical column was emulated using the Blue Gene/L supercomputer and the dynamics of the emulation reflected its biological counterpart.

Optogenetic silencing using halorhodopsin
(Han & Boyden, 2007)
- Ed Boyden continued developing optogenetic tools to manipulate neural activity. Along with Xue Han, he expressed a codon-optimized version of a bacterial halorhodopsin (along with the ChR2 protein) in neurons.
- Upon exposure to yellow light, halorhodopsin pumps chloride ions into the cell, hyperpolarizing the membrane and inhibiting neural activity.
- Using halorhodopsin and ChR2, neurons could be easily activated and inhibited using yellow and blue light respectively.
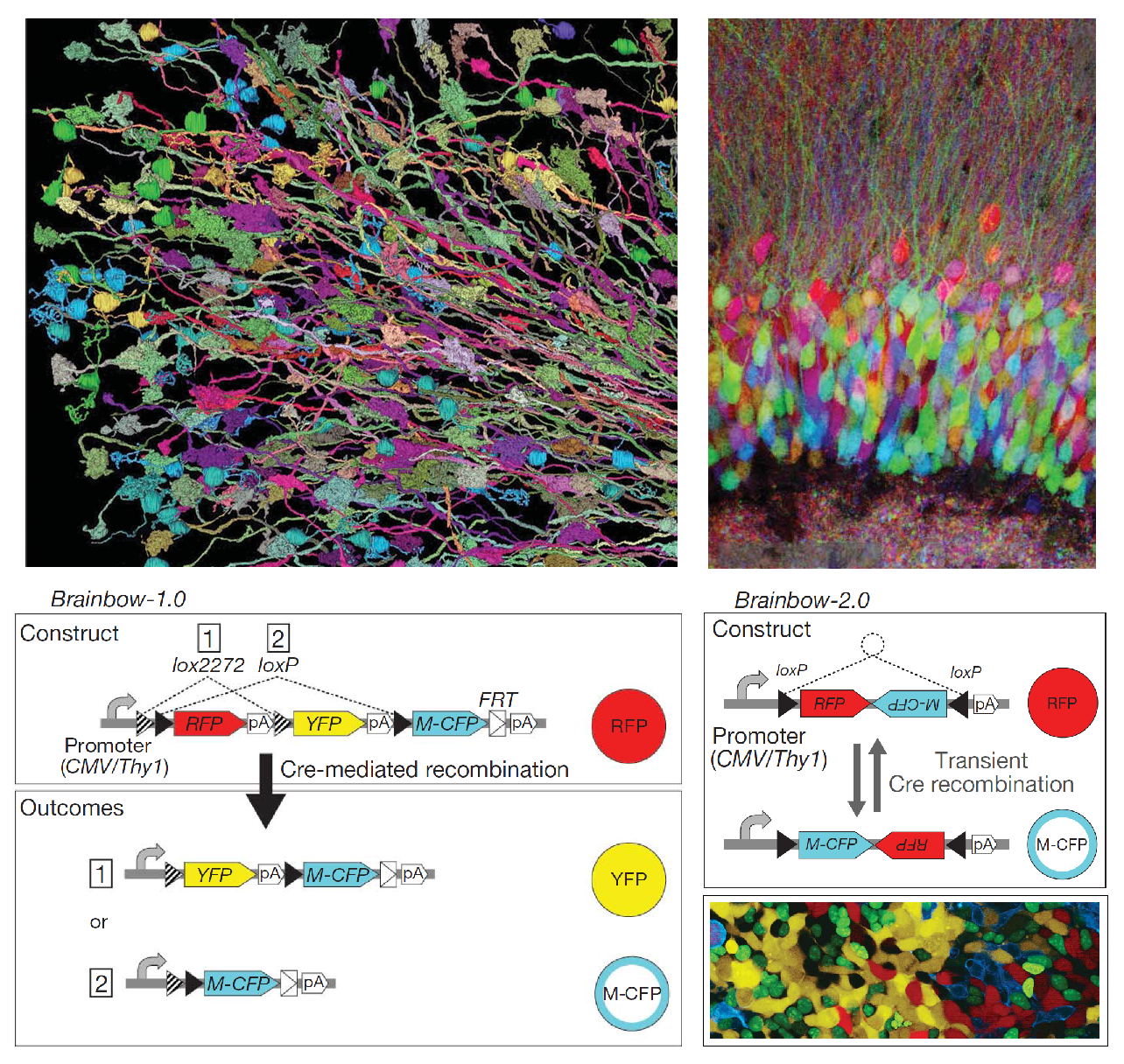
Brainbow
(Livet et al., 2007)
- Lichtman and colleagues used Cre/Lox recombination tools to create genes which express a randomized set of three or more differently-colored fluorescent proteins (XFPs) in a given neuron, labeling the neuron with a unique combination of colors. About ninety distinct colors were emitted across a population of genetically modified neurons.
- The detailed structures within neural tissue equipped with the Brainbow system can be imaged much more easily since neurons can be distinguished via color contrast.
- As a proof-of-concept, hundreds of synaptic contacts and axonal processes were reconstructed in a selected volume of the cerebellum. Several other neural structures were also imaged using Brainbow.
- The fluorescent proteins expressed by the Brainbow system are usable in vivo.
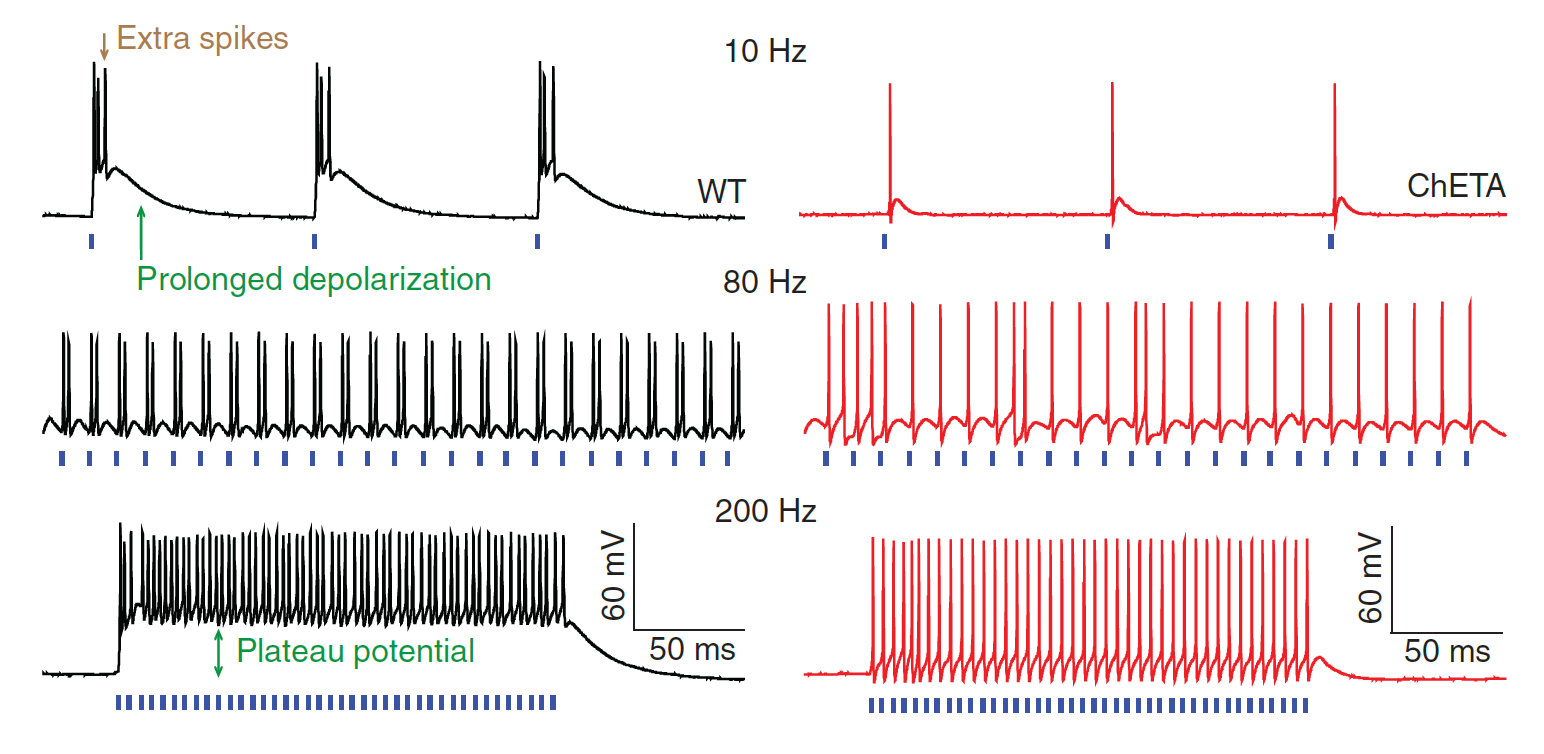
High temporal precision optogenetics
(Gunaydin et al., 2010)
- Karl Deisseroth, Peter Hegemann, and colleagues used protein engineering to improve the temporal resolution of optogenetic stimulation.
- Glutamic acid at position 123 in ChR2 was mutated to threonine, producing a new ion channel protein (dubbed ChETA).
- The ChETA protein allows for induction of spike trains with frequencies up to 200 Hz and greatly decreases the incidence of unintended spikes. Furthermore, ChETA eliminates plateau potentials (a phenomenon which interferes with precise control of neural activity).
Hippocampal prosthesis in rats
(Berger et al., 2012)
- Theodore Berger and his team developed an artificial replacement for neurons which transmit information from the CA3 region to the CA1 region of the hippocampus.
- This cognitive prosthesis employs recording and stimulation electrodes along with a multi-input multi-output (MIMO) model to encode the information in CA3 and transfer it to CA1.
- The hippocampal prosthesis was shown to restore and enhance memory in rats as evaluated by behavioral testing and brain imaging.
In vivo superresolution microscopy for neuroimaging
(Berning, Willig, Steffens, Dibaj, & Hell, 2012)
- Stefan Hell (2014 Nobel laureate in chemistry) developed stimulated emission depletion microscopy (STED), a type of superresolution fluorescence microscopy which allows imaging of synapses and dendritic spines.
- STED microscopy uses a torus-shaped de-excitation laser that interferes with the excitation laser to deplete fluorescence except in a very small spot. In this way, the diffraction limit is surpassed since the resulting light illuminates extremely small regions of the sample.
- Neurons in transgenic mice (equipped with glass-sealed holes in their skulls) were imaged using STED. Synapses and dendritic spines were observed up to fifteen nanometers below the surface of the brain tissue.
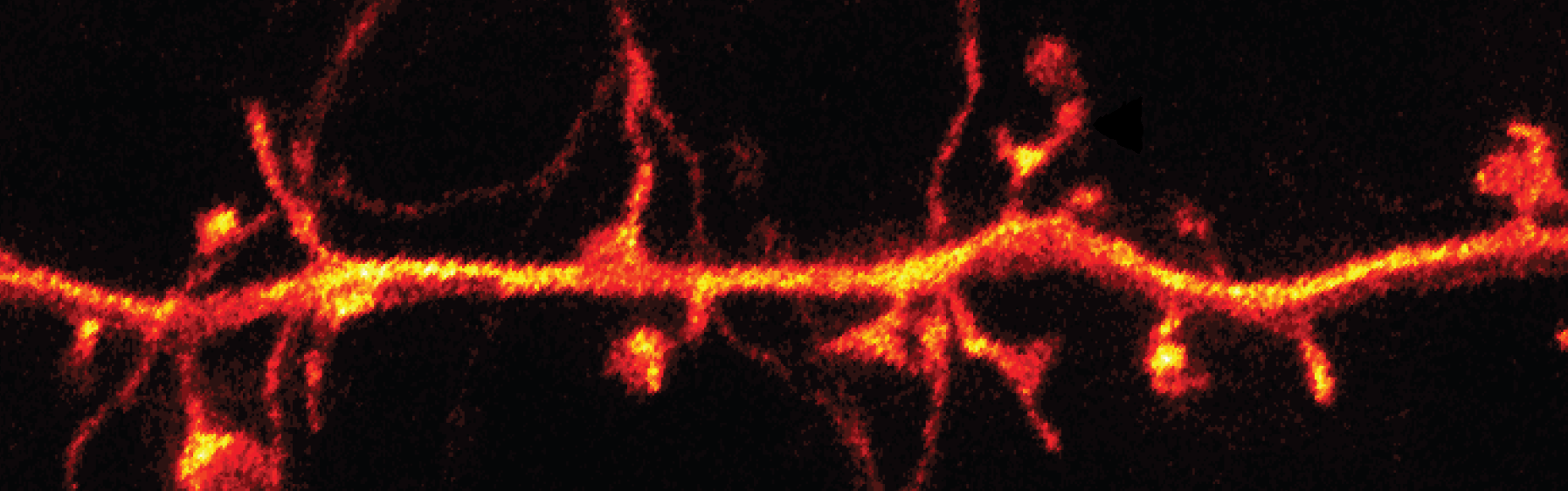
In vivo three-photon microscopy
(Horton et al., 2013)
- Multi-photon excitation uses pulsed lasers to excite fluorophores with two or more photons of light with long wavelengths. During the excitation, the photons undergo a nonlinear recombination process, yielding a single emitted photon with a much shorter wavelength. Because the excitation photons possess long wavelengths, they can penetrate tissue much more deeply than traditional microscopy allows.
- Horton and colleagues developed a three-photon excitation method to facilitate even deeper tissue penetration than the commonly used two-photon microscopic techniques.
- Since three photons were involved per excitation event, even longer excitation wavelengths (about 1,700 nm) were usable, allowing the construction of a 3-dimensional image stack that reached a depth of up to 1.4 mm within the living mouse brain.
- Blood vessels and RFP-labeled neurons were imaged using this approach. Furthermore, the depth was sufficient to enable imaging of neurons within the mouse hippocampus.

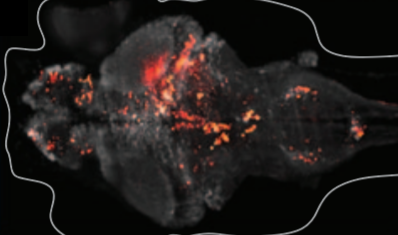
Whole-brain functional recording from larval zebrafish
(Ahrens, Orger, Robson, Li, & Keller, 2013)
- Laser-scanning light-sheet microscopy was used to volumetrically image the entire brains of larval zebrafish (an optically transparent organism).
- The genetically encoded calcium sensor GCaMP5G facilitated functional recording at single-cell resolution from about 80% of the total neurons in the larval zebrafish brains. Computational methods were used to distinguish between individual neurons.
- Populations of neurons that underwent correlated activity patterns were identifiedto show the technique’s utility for uncovering the dynamics of neural circuits. These populations included hindbrain neurons that were functionally linked to neural activity in the spinal cord and a population of neurons which showed coupled oscillations on the left and right halves.
Eyewire: crowdsourcing method for retina mapping
(Marx, 2013)
- The Eyewire project was created by Sebastian Seung’s research group. It is a crowdsourcing initiative for connectomic mapping within the retina towards uncovering neural circuits involved in visual processing.
- Laboratories first collect data via serial electron microscopy as well as functional data from two-photon microscopy.
- In the Eyewire game, images of tissue slices are provided to players who then help reconstruct neural morphologies and circuits by “coloring in” the parts of the images which correspond to cells and stacking many images on top of each other to generate 3D maps. Artificial intelligence tools help provide initial “best guesses” and guide the players, but the people ultimately perform the task of reconstruction.
- By November 2013, around 82,000 participants had played the game. Its popularity continues to grow.

The BRAIN Initiative
(“Fact Sheet: BRAIN Initiative,” 2013)
- The BRAIN Initiative (Brain Research through Advancing Innovative Technologies) provided neuroscientists with $110 million in governmental funding and $122 million in funding from private sources such as the Howard Hughes Medical Institute and the Allen Institute for Brain Science.
- The BRAIN Initiative focused on funding research which develops and utilizes new technologies for functional connectomics. It helped to accelerate research on tools for decoding the mechanisms of neural circuits in order to understand and treat mental illness, neurodegenerative diseases, and traumatic brain injury.
- The BRAIN Initiative emphasized collaboration between neuroscientists and physicists. It also pushed forward nanotechnology-based methods to image neural tissue, record from neurons, and otherwise collect neurobiological data.
The CLARITY method for making brains translucent
(Chung & Deisseroth, 2013)
- Karl Deisseroth and colleagues developed a method called CLARITY to make samples of neural tissue optically translucent without damaging the fine cellular structures in the tissue. Using CLARITY, entire mouse brains have been turned transparent.
- Mouse brains were infused with hydrogel monomers (acrylamide and bisacrylamide) as well as formaldehyde and some other compounds for facilitating crosslinking. Next, the hydrogel monomers were crosslinked by incubating the brains at 37°C. Lipids in the hydrogel-stabilized mouse brains were extracted using hydrophobic organic solvents and electrophoresis.
- CLARITY allows antibody labeling, fluorescence microscopy, and other optically-dependent techniques to be used for imaging entire brains. In addition, it renders the tissue permeable to macromolecules, which broadens the types of experimental techniques that these samples can undergo (i.e. macromolecule-based stains, etc.)

X-ray microtomography used to reconstruct Drosophila brain hemisphere
(Mizutani, Saiga, Takeuchi, Uesugi, & Suzuki, 2013)
- Mizutani and colleagues stained Drosophila brains with silver nitrate and tetrachloroaurate (a gold-containing compound), facilitating 3-dimensional imaging using X-ray microtomography at a voxel size of 220 × 328 × 314 nm.
- To generate the X-rays, a synchrotron source was used. It should be noted that synchrotron sources require large facilities to operate.
- Neuronal tracing was performed manually on the 3-dimensional X-ray images of the fly brain, a process which took about 1,700 person-hours. Some neuronal processes were too dense to be resolved, so they were “fused” into unified structures. Furthermore, some neuronal traces were fragmented and most of the cell bodies were not considered. This decreased the number of traces to one third of the estimated number of actual processes in the hemisphere.
- Mizutani’s investigation represents an early effort at large-scale connectomics that sets the stage for further initiatives as neuronal tracing, sample preparation, and X-ray microtomography technologies continue to improve.

Telepathic rats engineered using hippocampal prosthesis
(S. Deadwyler et al., 2013)
- Berger’s hippocampal prosthesis was implanted in pairs of rats. When “donor” rats were trained to perform a task, they developed neural representations (memories) which were recorded by their hippocampal prostheses.
- The donor rat memories were run through the MIMO model and transmitted to the stimulation electrodes of the hippocampal prostheses implanted in untrained “recipient” rats. After receiving the memories, the recipient rats showed significant improvements on the task that they had not been trained to perform.

Integrated Information Theory 3.0
(Oizumi, Albantakis, & Tononi, 2014)
- Integrated information theory (IIT) was originally proposed by Giulio Tononi in 2004. IIT is a quantitative theory of consciousness which may help explain the hard problem of consciousness.
- IIT begins by assuming the following phenomenological axioms; each experience is characterized by how it differs from other experiences, an experience cannot be reduced to interdependent parts, and the boundaries which distinguish individual experiences are describable as having defined “spatiotemporal grains.”
- From these phenomenological axioms and the assumption of causality, IIT identifies maximally irreducible conceptual structures (MICS) associated with individual experiences. MICS represent particular patterns of qualia that form unified percepts.
- IIT also outlines a mathematical measure of an experience’s quantity. This measure is called integrated information or ϕ.
Openworm
(Szigeti et al., 2014)
- The anatomical elegans connectome was originally mapped in 1976 by Albertson and Thomson. More data has since been collected on neurotransmitters, electrophysiology, cell morphology, and other characteristics.
- Szigeti, Larson, and their colleagues made an online platform for crowdsourcing research on elegans computational neuroscience, with the goal of completing an entire “simulated worm.”
- The group also released software called Geppetto, a program that allows users to manipulate both multicompartmental Hodgkin-Huxley models and highly efficient soft-body physics simulations (for modeling the worm’s electrophysiology and anatomy).
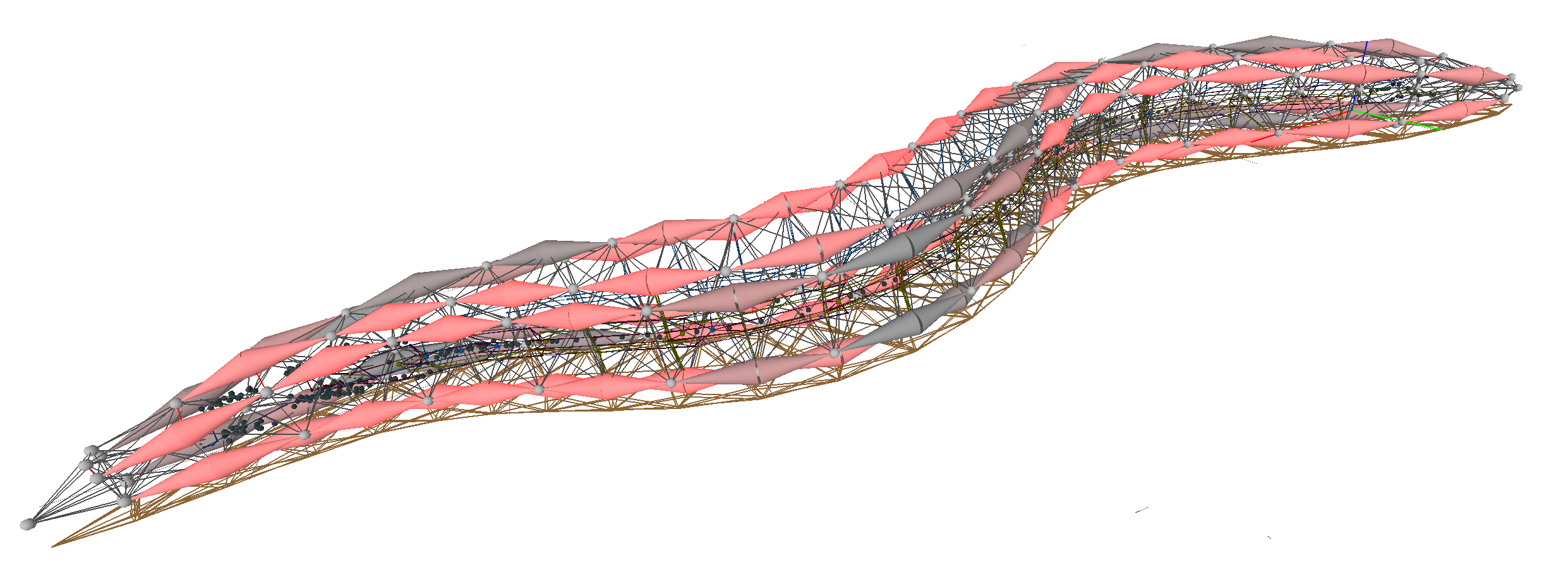
Expansion microscopy
(F. Chen, Tillberg, & Boyden, 2015)
- The Boyden group developed expansion microscopy, a method which enlarges neural tissue samples (including entire brains) with minimal structural distortions and so facilitates superior optical visualization of the scaled-up neural microanatomy. Furthermore, expansion microscopy greatly increases the optical translucency of treated samples.
- Expansion microscopy operates by infusing a swellable polymer network into brain tissue samples along with several chemical treatments to facilitate polymerization and crosslinking and then triggering expansion via dialysis in water. With 4.5-fold enlargement, expansion microscopy only distorts the tissue by about 1% (computed using a comparison between control superresolution microscopy of easily-resolvable cellular features and the expanded version).
- Before expansion, samples can express various fluorescent proteins to facilitate superresolution microscopy of the enlarged tissue once the process is complete. Furthermore, expanded tissue is highly amenable to fluorescent stains and antibody-based labels.

Japan’s Brain/MINDS project
(Okano, Miyawaki, & Kasai, 2015)
- In 2014, the Brain/MINDS (Brain Mapping by Integrated Neurotechnologies for Disease Studies) project was initiated to further neuroscientific understanding of the brain. This project received nearly $30 million in funding for its first year alone.
- Brain/MINDS focuses on studying the brain of the common marmoset (a non-human primate abundant in Japan), developing new technologies for brain mapping, and understanding the human brain with the goal of finding new treatments for brain diseases.
The TrueNorth chip from DARPA and IBM
(Akopyan et al., 2015)
- The TrueNorth neuromorphic computing chip was constructed and validated by DARPA and IBM. TrueNorth uses circuit modules which mimic neurons. Inputs to these fundamental circuit modules must overcome a threshold in order to trigger “firing.”
- The chip can emulate up to a million neurons with over 250 million synapses while requiring far less power than traditional computing devices.
Human Brain Project cortical mesocircuit reconstruction and simulation
(Markram et al., 2015)
- The Human Brain Project reconstructed a 0.29 mm3 region of rat cortical tissue including about 31,000 neurons and 37 million synapses based on morphological data, statistical connectivity rules (rather than exact connectivity), and other datasets. The cortical mesocircuit was emulated using the Blue Gene/Q supercomputer.
- This emulation was sufficiently accurate to reproduce emergent neurological processes and yield insights on the mechanisms of their computations.

Recording from C. elegans neurons reveals motor operations
(Kato et al., 2015)
- Live elegans worms were immobilized in microfluidic devices and the neurons in their head ganglia as well as some of their motor systems were imaged and recorded from using the calcium indicator GCaMP. As the C. elegans connectome is well-characterized, Kato and colleagues were able to determine the identities of most of the cells that underwent imaging (with the help of computational segmentation techniques).
- Principal component analysis was used to reduce the dimensionality of the neural activity datasets since over 100 neurons per worm were recorded from simultaneously.
- Next, phase space analysis was utilized to visualize the patterns formed by the recording data. Motor behaviors including dorsal turns, ventral turns, forward movements, and backward movements were found to correspond to specific sequences of neural events as uncovered by examining the patterns found in the phase plots. Further analyses revealed various insights about these brain dynamics and their relationship to motor actions.

Neural lace
(Liu et al., 2015)
- Charles Lieber’s group developed a syringe-injectable electronic mesh made of submicrometer-thick wiring for neural interfacing.
- The meshes were constructed using novel soft electronics for biocompatibility. Upon injection, the neural lace expands to cover and record from centimeter-scale regions of tissue.
- Neural lace may allow for “invasive” brain-computer interfaces to circumvent the need for surgical implantation. Lieber has continued to develop this technology towards clinical application.

BigNeuron initiative towards standardized neuronal morphology acquisition
(Peng et al., 2015)
- Because of the inconsistencies between neuronal reconstruction methods and lack of standardization found in neuronal morphology databases, BigNeuron was established as a community effort to improve the situation.
- BigNeuron tests as many automated neuronal reconstruction algorithms as possible using large-scale microscopy datasets (from several types of light microscopy). It uses the Vaa3D neuronal reconstruction software as a central platform. Reconstruction algorithms are added to Vaa3D as plugins. These computational tests are performed on supercomputers.
- BigNeuron aims to create a superior community-oriented neuronal morphology database, a set of greatly improved tools for neuronal reconstruction, a standardized protocol for future neuronal reconstructions, and a library of morphological feature definitions to facilitate classification.
Human telepathy during a 20 questions game
(Stocco et al., 2015)
- Using an interactive question-and-answer setup, Stocco and colleagues demonstrated real-time telepathic communication between pairs of individuals via EEG and transcranial magnetic stimulation. Five pairs of participants played games of 20 questions and attempted to identify unknown objects.
- EEG data were recorded from the respondent, computationally processed, and transmitted as transcranial magnetic stimulation signals into the mind (occipital lobe stimulation) of a respondent. The respondent’s answers were translated into higher-intensity transcranial magnetic stimulation pulses corresponding to “yes” answers or lower-intensity transcranial magnetic stimulation pulses corresponding to “no” answers.
- When compared to control trials in which sham interfaces were used, the people using the brain-brain interfaces were significantly more successful at playing 20 questions games.
Expansion FISH
(F. Chen et al., 2016)
- Boyden, Chen, Marblestone, Church, and colleagues combined fluorescent in situ hybridization (FISH) with expansion microscopy to image the spatial localization of RNA in neural tissue.
- The group developed a chemical linker to covalently attach intracellular RNA to the infused polymer network used in expansion microscopy. This allowed for RNAs to maintain their relative spatial locations within each cell post-expansion.
- After the tissue was enlarged, FISH was used to fluorescently label targeted RNA molecules. In this way, RNA localization was more effectively resolved.
- As a proof-of-concept, expansion FISH was used to reveal the nanoscale distribution of long noncoding RNAs in nuclei as well as the locations of RNAs within dendritic spines.

Neural dust
(Seo et al., 2016)
- Michel Maharbiz’s group invented implantable, ~ 1 mm biosensors for wireless neural recording and tested them in rats.
- This neural dust could be miniaturized to less than 0.5 mm or even to microscale dimensions using customized electronic components.
- Neural dust motes consist of two recording electrodes, a transistor, and a piezoelectric crystal.
- The neural dust received external power from ultrasound. Neural signals were recorded by measuring disruptions to the piezoelectric crystal’s reflection of the ultrasound waves. Signal processing mathematics allowed precise detection of activity.
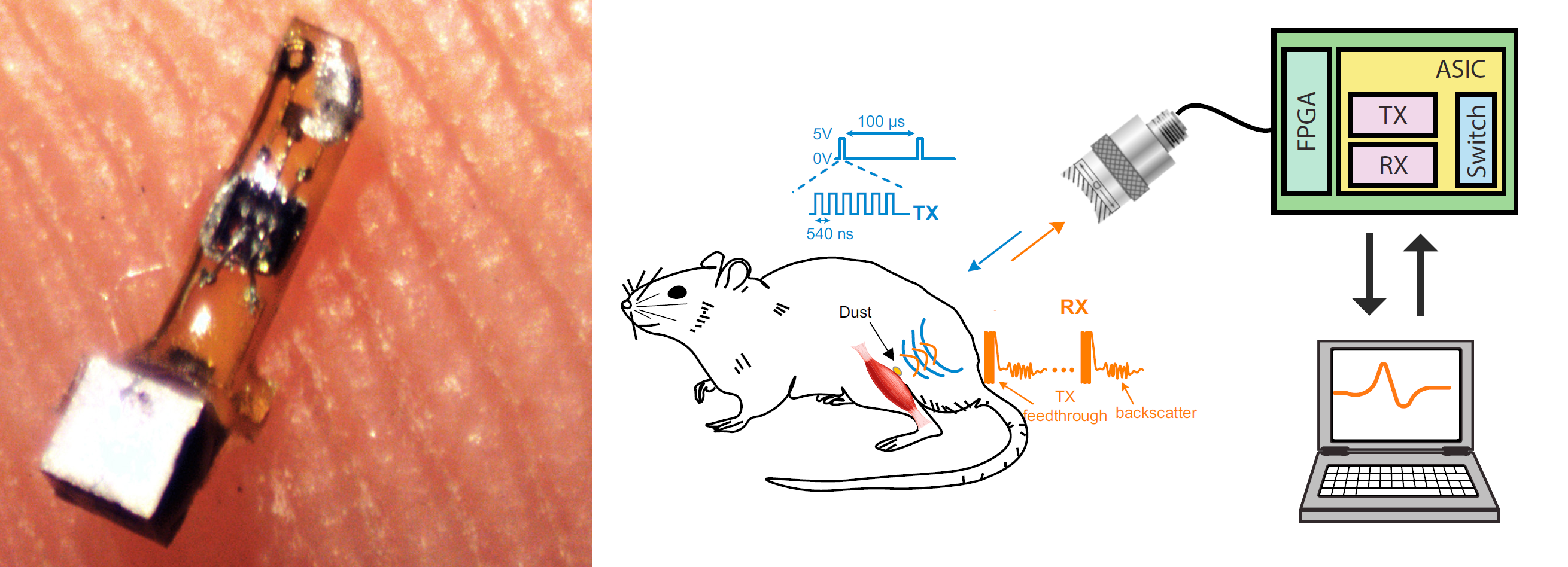
The China Brain Project
(Poo et al., 2016)
- The China Brain Project was launched to help understand the neural mechanisms of cognition, develop brain research technology platforms, develop preventative and diagnostic interventions for brain disorders, and to improve brain-inspired artificial intelligence technologies.
- This project will be take place from 2016 until 2030 with the goal of completing mesoscopic brain circuit maps.
- China’s population of non-human primates and preexisting non-human primate research facilities give the China Brain Project an advantage. The project will focus on studying rhesus macaques.
Somatosensory cortex stimulation for spinal cord injuries
(Flesher et al., 2016)
- Gaunt, Flesher, and colleagues found that microstimulation of the primary somatosensory cortex (S1) partially restored tactile sensations to a patient with a spinal cord injury.
- Electrode arrays were implanted into the S1 regions of a patient with a spinal cord injury. The array performed intracortical microstimulation over a period of six months.
- The patient reported locations and perceptual qualities of the sensations elicited by microstimulation. The patient did not experience pain or “pins and needles” from any of the stimulus trains. Overall, 93% of the stimulus trains were reported as “possibly natural.”
- Results from this study might be used to engineer upper-limb neuroprostheses which provide somatosensory feedback.

Simulation of rat CA1 region
(Bezaire, Raikov, Burk, Vyas, & Soltesz, 2016)
- Detailed computational models of 338,740 neurons (including pyramidal cells and various types of interneurons) were equipped with connectivity patterns based on data from the biological CA1 region. External inputs were also estimated using biological data and incorporated into the simulation. It is important to note that these connectivity patterns described the typical convergence and divergence of neurites to and from particular cell types rather than explicitly representing the exact connections found in the biological rat.
- Each neuron was simulated using a multicompartmental Hodgkin-Huxley-type model with its morphological structure based on biological data from the given cell type. Furthermore, different cell types received different numbers of presynaptic terminals at specified distances from the soma. In total, over five billion synapses were present within the CA1 model.
- The simulation was implemented on several different supercomputers. Due to the model’s complexity, a four second simulation took about four hours to complete.
- As with the biological CA1 region, the simulation gave rise to gamma oscillations and theta oscillations as well as other biologically consistent phenomena. In addition, parvalbumin-expressing interneurons and neurogliaform cells were identified as drivers of the theta oscillations, demonstrating the utility of detailed neuronal simulations for uncovering biological insights.

UltraTracer enhances existing neuronal tracing software
(Peng et al., 2017)
- UltraTracer is an algorithm that can improve the efficiency of existing neuronal tracing software for handling large datasets while maintaining accuracy.
- Datasets with hundreds of billions of voxels were utilized to test UltraTracer. Ten existing tracing algorithms were augmented.
- For most of the existing algorithms, the performance improvements were around 3-6 times, though a few showed improvements of 10-30 times. Even when using computers with smaller memory, UltraTracer was consistently able to enhance conventional software.
- UltraTracer was made opensource and is available as a plugin for the Vaa3D tracing software suite.
Whole-brain electron microscopy in larval zebrafish
(Hildebrand et al., 2017)
- Serial electron microscopy facilitated imaging of the entire brain of a larval zebrafish at 5.5 days post-fertilization.
- Neuronal tracing software (a modified version of the CATMAID software) was used to reconstruct all the myelinated axons found in the larval zebrafish brain.
- The reconstructed dataset included 2,589 myelinated axon segments along with some of the associated soma and dendrites. It should be noted that only 834 of the myelinated axons were successfully traced back to their cell bodies.

Hippocampal prosthesis in monkeys
(S. A. Deadwyler et al., 2017)
- Theodore Berger continued developing his cognitive prosthesis and tested it in Rhesus Macaques.
- As with the rats, monkeys with the implant showed substantially improved performance on memory tasks.
The $100 billion Softbank Vision Fund
(Lomas, 2017)
- Masayoshi Son, the CEO of Softbank (a Japanese telecommunications corporation), announced a plan to raise $100 billion in venture capital to invest in artificial intelligence. This plan involved partnering with multiple large companies in order to raise this enormous amount of capital.
- By the end of 2017, the Vision Fund successfully reached its $100 billion goal. Masayoshi Son has since announced further plans to continue raising money with a new goal of over $800 billion.
- Masayoshi Son’s reason for these massive investments is the Technological Singularity. He agrees with Kurzweil that the Singularity will likely occur at around 2045 and he hopes to help bring the Singularity to fruition. Though Son is aware of the risks posed by artificial superintelligence, he feels that superintelligent AI’s potential to tackle some of humanity’s greatest challenges (such as climate change and the threat of nuclear war) outweighs those risks.
Bryan Johnson launches Kernel
(Regalado, 2017)
- Entrepreneur Bryan Johnson invested $100 million to start Kernel, a neurotechnology company.
- Kernel plans to develop implants that allow for recording and stimulation of large numbers of neurons at once. The company’s initial goal is to develop treatments for mental illnesses and neurodegenerative diseases. Its long-term goal is to enhance human intelligence.
- Kernel originally partnered with Theodore Berger and intended to utilize his hippocampal prosthesis. Unfortunately, Berger and Kernel parted ways after about six months because Berger’s vision was reportedly too long-range to support a financially viable company (at least for now).
- Kernel was originally a company called Kendall Research Systems. This company was started by a former member of the Boyden lab. In total, four members of Kernel’s team are former Boyden lab members.
Elon Musk launches NeuraLink
(Etherington, 2017)
- Elon Musk (CEO of Tesla, SpaceX, and a number of other successful companies) initiated a neuroengineering venture called NeuraLink.
- NeuraLink will begin by developing brain-computer interfaces (BCIs) for clinical applications, but the ultimate goal of the company is to enhance human cognitive abilities in order to keep up with artificial intelligence.
- Though many of the details around NeuraLink’s research are not yet open to the public, it has been rumored that injectable electronics similar to Lieber’s neural lace might be involved.
Facebook announces effort to build brain-computer interfaces
(Constine, 2017)
- Facebook revealed research on constructing non-invasive brain-computer interfaces (BCIs) at a company-run conference in 2017. The initiative is run by Regina Dugan, Facebook’s head of R&D at division building 8.
- Facebook’s researchers are working on a non-invasive BCI which may eventually enable users to type one hundred words per minute with their thoughts alone. This effort builds on past investigations which have been used to help paralyzed patients.
- The building 8 group is also developing a wearable device for “skin hearing.” Using just a series of vibrating actuators which mimic the cochlea, test subjects have so far been able to recognize up to nine words. Facebook intends to vastly expand this device’s capabilities.
DARPA funds research to develop improved brain-computer interfaces
(Hatmaker, 2017)
- The U.S. government agency DARPA awarded $65 million in total funding to six research groups.
- The recipients of this grant included five academic laboratories (headed by Arto Nurmikko, Ken Shepard, Jose-Alain Sahel and Serge Picaud, Vicent Pieribone, and Ehud Isacoff) and one small company called Paradromics Inc.
- DARPA’s goal for this initiative is to develop a nickel-sized bidirectional brain-computer interface (BCI) which can record from and stimulate up to one million individual neurons at once.
Human Brain Project analyzes brain computations using algebraic topology
(Reimann et al., 2017)
- Investigators at the Human Brain Project utilized algebraic topology to analyze the reconstructed ~ 31,000 neuron cortical microcircuit from their earlier work.
- The analysis involved representing the cortical network as a digraph, finding directed cliques (complete directed subgraphs belonging to a digraph), and determining the net directionality of information flow (by computing the sum of the squares of the differences between in-degree and out-degree for all the neurons in a clique). In algebraic topology, directed cliques of n neurons are called directed simplices of dimension n-1.
- Vast numbers of high-dimensional directed cliques were found in the cortical microcircuit (as compared to null models and other controls). Spike correlations between pairs of neurons within a clique were found to increase with the clique’s dimension and with the proximity of the neurons to the clique’s sink. Furthermore, topological metrics allowed insights into the flow of neural information among multiple cliques.
- Experimental patch-clamp data supported the significance of the findings. In addition, similar patterns were found within the elegans connectome, suggesting that the results may generalize to nervous systems across species.

Early testing of hippocampal prosthesis algorithm in humans
(Song, She, Hampson, Deadwyler, & Berger, 2017)
- Dong Song (who was working alongside Berger) tested the MIMO algorithm on human epilepsy patients using implanted recording and stimulation electrodes. The full hippocampal prosthesis was not implanted, but the electrodes acted similarly, though in a temporary capacity. Although only two patients were tested in this study, many trials were performed to compensate for the small sample size.
- Hippocampal spike trains from individual cells in CA1 and CA3 were recorded from the patients during a delayed match-to-sample task. The patients were shown various images while neural activity data were recorded by the electrodes and processed by the MIMO model. The patients were then asked to recall which image they had been shown previously by picking it from a group of “distractor” images. Memories encoded by the MIMO model were used to stimulate hippocampal cells during the recall phase.
- In comparison to controls in which the same two epilepsy patients were not assisted by the algorithm and stimulation, the experimental trials demonstrated a significant increase in successful pattern matching.
Brain imaging factory in China
(Cyranoski, 2017)
- Qingming Luo started the HUST-Suzhou Institute for Brainsmatics, a brain imaging “factory.” Each of the numerous machines in Luo’s facility performs automated processing and imaging of tissue samples. The devices make ultrathin slices of brain tissue using diamond blades, treat the samples with fluorescent stains or other contrast-enhancing chemicals, and image then using fluorescence microscopy.
- The institute has already demonstrated its potential by mapping the morphology of a previously unknown neuron which “wraps around” the entire mouse brain.

Automated patch-clamp robot for in vivo neural recording
(Suk et al., 2017)
- Ed Boyden and colleagues developed a robotic system to automate patch-clamp recordings from individual neurons. The robot was tested in vivo using mice and achieved a data collection yield similar to that of skilled human experimenters.
- By continuously imaging neural tissue using two-photon microscopy, the robot can adapt to a target cell’s movement and shift the pipette to compensate. This adaptation is facilitated by a novel algorithm called an imagepatching algorithm. As the pipette approaches its target, the algorithm adjusts the pipette’s trajectory based on the real-time two-photon microscopy.
- The robot can be used in vivo so long as the target cells express a fluorescent marker or otherwise fluoresce corresponding to their size and position.

Genome editing in the mammalian brain
(Nishiyama, Mikuni, & Yasuda, 2017)
- Precise genome editing in the brain has historically been challenging because most neurons are postmitotic (non-dividing) and the postmitotic state prevents homology-directed repair (HDR) from occurring. HDR is a mechanism of DNA repair which allows for targeted insertions of DNA fragments with overhangs homologous to the region of interest (by contrast, non-homologous end-joining is highly unpredictable).
- Nishiyama, Mikuni, and Yasuda developed a technique which allows genome editing in postmitotic mammalian neurons using adeno-associated viruses (AAVs) and CRISPR-Cas9.
- The AAVs delivered ssDNA sequences encoding a single guide RNA (sgRNA) and an insert. Inserts encoding a hemagglutinin tag (HA) and inserts encoding EGFP were both tested. Cas9 was encoded endogenously by transgenic host cells and in transgenic host animals.
- The technique achieved precise genome editing in vitro and in vivo with a low rate of off-target effects. Inserts did not cause deletion of nearby endogenous sequences for 98.1% of infected neurons.
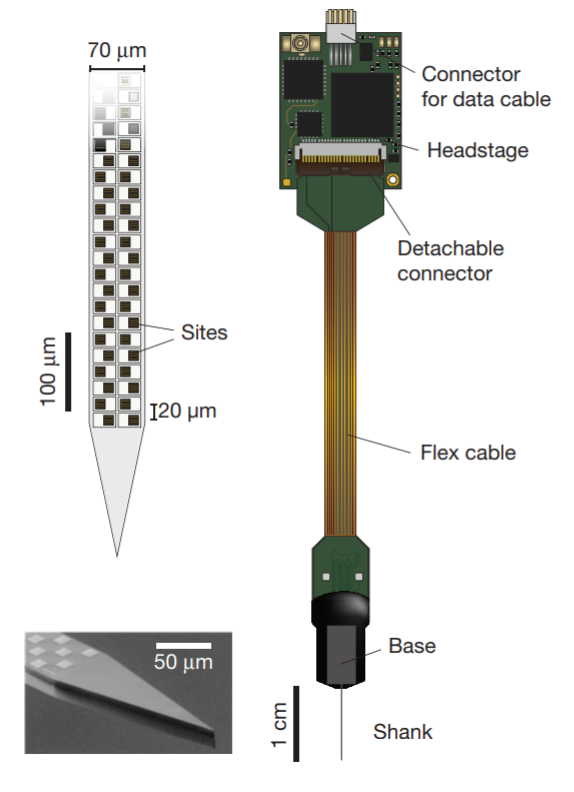
 Neuropixels probe
Neuropixels probe
(Jun et al., 2017)
- Jun and colleagues created the Neuropixels probe to facilitate simultaneous recording from hundreds of individual neurons with high spatiotemporal resolution. Previous extracellular probes were only able to record from a few dozen individual neurons.
- The Neuropixels recording shank is one centimeter long and includes 384 recording channels. Due to the small size of the accompanying apparatus (a 6×9 mm base and a data transmission cable), it enables high-throughput recording in freely moving animals. Because the shank is quite long, Neuropixels can record from multiple brain regions at once.
- Voltage signals are processed directly on the base of the Neuropixels apparatus, allowing for noise-free data transmission along the cable for further analysis.
EEG-based facial image reconstruction
(Nemrodov, Niemeier, Patel, & Nestor, 2018)
- EEG data associated with viewing images of faces was collected and used to determine the neural correlates of facial processing. In this way, the images were computationally reconstructed in a fashion resembling “mind reading.”
- It should be noted that the images reconstructed using data taken from multiple people were more accurate than the images reconstructed using single individuals. Nonetheless, the single individual data still yielded statistically significant accuracy.
- In addition to reconstructing the images themselves, the process gave insights on the cognitive steps involved in perceiving faces.

Near-infrared light and upconversion nanoparticles for optogenetic stimulation
(S. Chen et al., 2018)
- Upconversion nanoparticles absorb two or more low-energy photons and emit a higher energy photon. For instance, multiple near-infrared photons can be converted into a single visible spectrum photon.
- Shuo Chen and colleagues injected upconversion nanoparticles into the brains of mice and used them to convert externally applied near-infrared (NIR) light into visible light within the brain tissue. In this way, optogenetic stimulation was performed without the need for surgical implantation of fiber optics or similarly invasive procedures.
- The authors demonstrated stimulation via upconversion of NIR to blue light (to activate ChR2) and inhibition via upconversion of NIR to green light (to activate a rhodopsin called Arch).
- As a proof-of-concept, this technology was used to alter the behavior of the mice by activating hippocampally-encoded fear memories.

Map of all neuronal cell bodies within mouse brain
(Murakami et al., 2018)
- Ueda, Murakami, and colleagues combined methods from expansion microscopy and CLARITY to develop a protocol called CUBIC-X which both expands and clears entire brains. Light-sheet fluorescence microscopy was used to image the treated brains and a novel algorithm was developed to detect individual nuclei.
- Although expansion microscopy causes some increased tissue transparency on its own, CUBIC-X greatly improved this property in the enlarged tissues, facilitating more detailed whole-brain imaging.
- Using CUBIC-X, the spatial locations of all the cell bodies (but not dendrites, axons, or synapses) within the mouse brain were mapped. This process was performed upon several adult mouse brains as well as several developing mouse brains to allow for comparative analysis.
- The authors made the spatial atlas publicly available in order to facilitate global cooperation towards annotating connectivity among the neural cell bodies within the atlas.
Clinical testing of hippocampal prosthesis algorithm in humans
(Hampson et al., 2018)
- Further clinical tests of Berger’s hippocampal prosthesis were performed. Twenty-one patients took part in the experiments. Seventeen patients underwent CA3 recording so as to facilitate training and optimization of the MIMO model. Eight patients received CA1 stimulation so as to improve their memories.
- Electrodes with the ability to record from single neurons (10-24 single-neuron recording sites) and via EEG (4-6 EEG recording sites) were implanted such that recording and stimulation could occur at CA3 and CA1 respectively.
- Patients performed behavioral memory tasks. Both short-term and long-term memory showed an average improvement of 35% across the patients who underwent stimulation.
Precise optogenetic manipulation of fifty neurons
(Mardinly et al., 2018)
- Mardinly and colleagues engineered a novel excitatory optogenetic ion channel called ST-ChroME and a novel inhibitory optogenetic ion channel called IRES-ST-eGtACR1. The channels were localized to the somas of host neurons and generated stronger photocurrents over shorter timescales than previously existing opsins, allowing for powerful and precise optogenetic stimulation and inhibition.
- 3D-SHOT is an optical technique in which light is tuned by a device called a spatial light modulator along with several other optical components. Using 3D-SHOT, light was precisely projected upon targeted neurons within a volume of 550×550×100 μm3.
- By combining novel optogenetic ion channels and the 3D-SHOT technique, complex patterns of neural activity were created in vivo with high spatial and temporal precision.
- Simultaneously, calcium imaging allowed measurement of the induced neural activity. More custom optoelectronic components helped avoid optical crosstalk of the fluorescent calcium markers with the photostimulating laser.

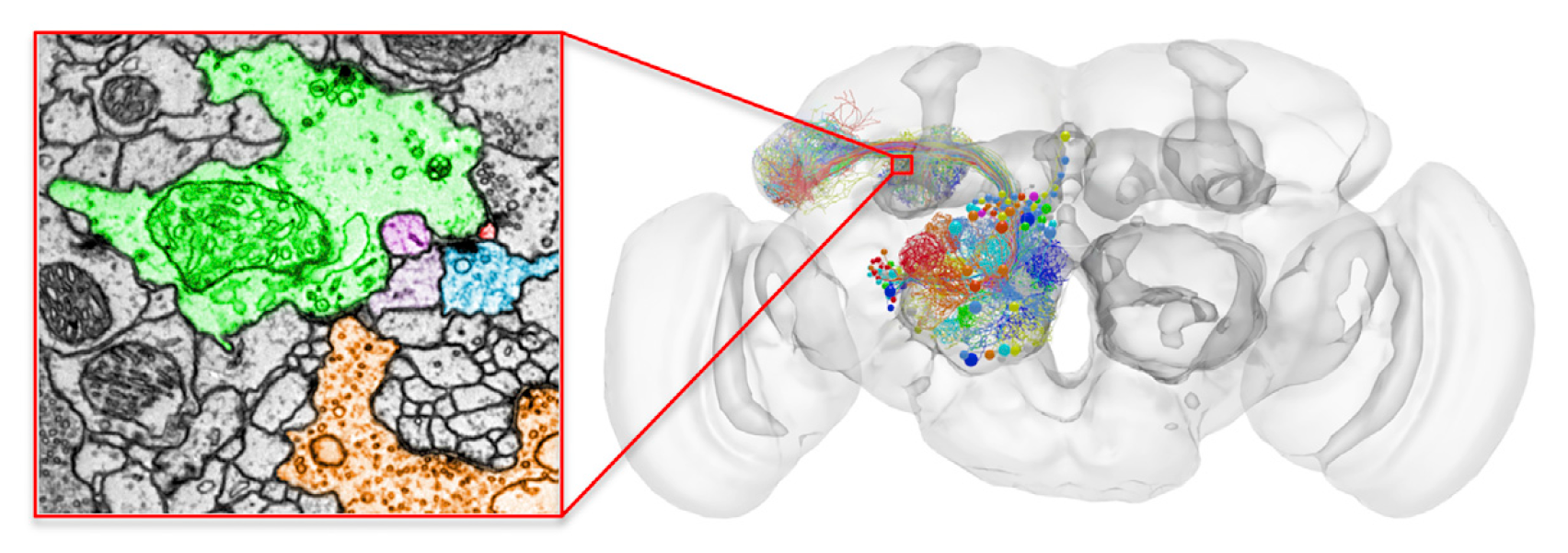
Whole-brain Drosophila connectome data acquired via serial electron microscopy
(Zheng et al., 2018)
- Zheng, Bock, and colleagues collected serial electron microscopy data on the entire adult Drosophila connectome, providing the data necessary to reconstruct a complete structural map of the fly’s brain at the resolution of individual synapses, dendritic spines, and axonal processes.
- The data are in the form of 7050 transmission electron microscopy images (187500 x 87500 pixels and 16 GB per image), each representing a 40nm-thin slice of the fly’s brain. In total the dataset requires 106 TB of storage.
- Although much of the the data still must be processed to reconstruct a 3-dimensional map of the Drosophila brain, the authors did create 3-dimensional reconstructions of selected areas in the olfactory pathway of the fly. In doing so, they discovered a new cell type as well as several other previously unrealized insights about the organization of Drosophila’s olfactory biology.
Human telepathy using BrainNet
(Jiang et al., 2018)
- EEG recordings were taken from two individuals (termed senders) while they played a Tetris-like game. Next, the recordings were converted into transcranial magnetic stimulation signals that acted to provide a third individual (called a receiver) with the necessary information to make decisions in the game without seeing the screen. The occipital cortex was stimulated. Fifteen people (five groups of three) took part in the study.
- To convey their information, the senders were told to focus upon either a higher or a lower intensity light corresponding to commands within the game (the two lights were placed on different sides of the computer screen). In the receiver’s mind, this translated to perceiving a flash of light. The receiver was able to distinguish the intensities and implement the correct command within the game.
- Using only the telepathically provided stimulation, the receiver made the correct game-playing decisions 81% of the time.

Transcriptomic cell type classification across mouse neocortex
(Tasic et al., 2018)
- Single-cell RNA sequencing was used to characterize gene expression across 23,822 cells from the primary visual cortex and the anterior lateral motor cortex of mice.
- Using dimensionality reduction and clustering methods, the resulting data were used to classify the neurons into 133 transcriptomic cell types.
- Injections of adeno associated viruses (engineered to express fluorescent markers) facilitated retrograde tracing of neuronal projections within a subset of the sequenced cells. In this way, correspondences between projection patterns and transcriptomic identities were established.
References
Ahrens, M. B., Orger, M. B., Robson, D. N., Li, J. M., & Keller, P. J. (2013). Whole-brain functional imaging at cellular resolution using light-sheet microscopy. Nature Methods, 10, 413. Retrieved from https://doi.org/10.1038/nmeth.2434
Akopyan, F., Sawada, J., Cassidy, A., Alvarez-Icaza, R., Arthur, J., Merolla, P., … Modha, D. S. (2015). TrueNorth: Design and Tool Flow of a 65 mW 1 Million Neuron Programmable Neurosynaptic Chip. IEEE Transactions on Computer-Aided Design of Integrated Circuits and Systems, 34(10), 1537–1557. https://doi.org/10.1109/TCAD.2015.2474396
Berger, T. W., Song, D., Chan, R. H. M., Marmarelis, V. Z., LaCoss, J., Wills, J., … Granacki, J. J. (2012). A Hippocampal Cognitive Prosthesis: Multi-Input, Multi-Output Nonlinear Modeling and VLSI Implementation. IEEE Transactions on Neural Systems and Rehabilitation Engineering, 20(2), 198–211. https://doi.org/10.1109/TNSRE.2012.2189133
Berning, S., Willig, K. I., Steffens, H., Dibaj, P., & Hell, S. W. (2012). Nanoscopy in a Living Mouse Brain. Science, 335(6068), 551 LP-551. Retrieved from http://science.sciencemag.org/content/335/6068/551.abstract
Bezaire, M. J., Raikov, I., Burk, K., Vyas, D., & Soltesz, I. (2016). Interneuronal mechanisms of hippocampal theta oscillations in a full-scale model of the rodent CA1 circuit. ELife, 5, e18566. https://doi.org/10.7554/eLife.18566
Boyden, E. S., Zhang, F., Bamberg, E., Nagel, G., & Deisseroth, K. (2005). Millisecond-timescale, genetically targeted optical control of neural activity. Nature Neuroscience, 8, 1263. Retrieved from http://dx.doi.org/10.1038/nn1525
Chen, F., Tillberg, P. W., & Boyden, E. S. (2015). Expansion microscopy. Science, 347(6221), 543 LP-548. Retrieved from http://science.sciencemag.org/content/347/6221/543.abstract
Chen, F., Wassie, A. T., Cote, A. J., Sinha, A., Alon, S., Asano, S., … Boyden, E. S. (2016). Nanoscale imaging of RNA with expansion microscopy. Nature Methods, 13, 679. Retrieved from http://dx.doi.org/10.1038/nmeth.3899
Chen, S., Weitemier, A. Z., Zeng, X., He, L., Wang, X., Tao, Y., … McHugh, T. J. (2018). Near-infrared deep brain stimulation via upconversion nanoparticle–mediated optogenetics. Science, 359(6376), 679 LP-684. Retrieved from http://science.sciencemag.org/content/359/6376/679.abstract
Chung, K., & Deisseroth, K. (2013). CLARITY for mapping the nervous system. Nature Methods, 10, 508. Retrieved from http://dx.doi.org/10.1038/nmeth.2481
Constine, J. (2017). Facebook is building brain-computer interfaces for typing and skin-hearing. TechCrunch. Retrieved from https://techcrunch.com/2017/04/19/facebook-brain-interface/
Cyranoski, D. (2017). China launches brain-imaging factory. Nature, 548(7667), 268–269. https://doi.org/10.1038/548268a
Deadwyler, S. A., Hampson, R. E., Song, D., Opris, I., Gerhardt, G. A., Marmarelis, V. Z., & Berger, T. W. (2017). A cognitive prosthesis for memory facilitation by closed-loop functional ensemble stimulation of hippocampal neurons in primate brain. Experimental Neurology, 287, 452–460. https://doi.org/https://doi.org/10.1016/j.expneurol.2016.05.031
Deadwyler, S., Hampson, R., Sweat, A., Song, D., Chan, R., Opris, I., … Berger, T. (2013). Donor/recipient enhancement of memory in rat hippocampus. Frontiers in Systems Neuroscience. Retrieved from https://www.frontiersin.org/article/10.3389/fnsys.2013.00120
Etherington, D. (2017). Elon Musk’s Neuralink wants to boost the brain to keep up with AI. TechCrunch. Retrieved from techcrunch.com/2017/03/27/elon-musks-neuralink-wants-to-boost-the-brain-to-keep-up-with-ai/
Fact Sheet: BRAIN Initiative. (2013). Retrieved from https://obamawhitehouse.archives.gov/the-press-office/2013/04/02/fact-sheet-brain-initiative
Flesher, S. N., Collinger, J. L., Foldes, S. T., Weiss, J. M., Downey, J. E., Tyler-Kabara, E. C., … Gaunt, R. A. (2016). Intracortical microstimulation of human somatosensory cortex. Science Translational Medicine. Retrieved from http://stm.sciencemag.org/content/early/2016/10/12/scitranslmed.aaf8083.abstract
Gunaydin, L. A., Yizhar, O., Berndt, A., Sohal, V. S., Deisseroth, K., & Hegemann, P. (2010). Ultrafast optogenetic control. Nature Neuroscience, 13, 387. Retrieved from http://dx.doi.org/10.1038/nn.2495
Hampson, R. E., Song, D., Robinson, B. S., Fetterhoff, D., Dakos, A. S., Roeder, B. M., … Deadwyler, S. A. (2018). Developing a hippocampal neural prosthetic to facilitate human memory encoding and recall. Journal of Neural Engineering, 15(3), 36014. https://doi.org/10.1088/1741-2552/aaaed7
Han, X., & Boyden, E. S. (2007). Multiple-Color Optical Activation, Silencing, and Desynchronization of Neural Activity, with Single-Spike Temporal Resolution. PLOS ONE, 2(3), e299. Retrieved from https://doi.org/10.1371/journal.pone.0000299
Hatmaker, T. (2017). DARPA awards $65 million to develop the perfect, tiny two-way brain-computer interface. TechCrunch. Retrieved from techcrunch.com/2017/07/10/darpa-nesd-grants-paradromics/
Hildebrand, D. G. C., Cicconet, M., Torres, R. M., Choi, W., Quan, T. M., Moon, J., … Engert, F. (2017). Whole-brain serial-section electron microscopy in larval zebrafish. Nature, 545, 345. Retrieved from https://doi.org/10.1038/nature22356
Horton, N. G., Wang, K., Kobat, D., Clark, C. G., Wise, F. W., Schaffer, C. B., & Xu, C. (2013). In vivo three-photon microscopy of subcortical structures within an intact mouse brain. Nature Photonics, 7, 205. Retrieved from https://doi.org/10.1038/nphoton.2012.336
Jiang, L., Stocco, A., Losey, D. M., Abernethy, J. A., Prat, C. S., & Rao, R. P. N. (2018). BrainNet: a multi-person brain-to-brain interface for direct collaboration between brains. ArXiv Preprint ArXiv:1809.08632.
Jun, J. J., Steinmetz, N. A., Siegle, J. H., Denman, D. J., Bauza, M., Barbarits, B., … Harris, T. D. (2017). Fully integrated silicon probes for high-density recording of neural activity. Nature, 551, 232. Retrieved from https://doi.org/10.1038/nature24636
Kato, S., Kaplan, H. S., Schrödel, T., Skora, S., Lindsay, T. H., Yemini, E., … Zimmer, M. (2015). Global Brain Dynamics Embed the Motor Command Sequence of Caenorhabditis elegans. Cell, 163(3), 656–669. https://doi.org/10.1016/j.cell.2015.09.034
Liu, J., Fu, T.-M., Cheng, Z., Hong, G., Zhou, T., Jin, L., … Lieber, C. M. (2015). Syringe-injectable electronics. Nature Nanotechnology, 10, 629. Retrieved from http://dx.doi.org/10.1038/nnano.2015.115
Livet, J., Weissman, T. A., Kang, H., Draft, R. W., Lu, J., Bennis, R. A., … Lichtman, J. W. (2007). Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature, 450, 56. Retrieved from http://dx.doi.org/10.1038/nature06293
Lomas, N. (2017). Superintelligent AI explains Softbank’s push to raise a $100BN Vision Fund. TechCrunch. Retrieved from https://techcrunch.com/2017/02/27/superintelligent-ai-explains-softbanks-push-to-raise-a-100bn-vision-fund/
Mardinly, A. R., Oldenburg, I. A., Pégard, N. C., Sridharan, S., Lyall, E. H., Chesnov, K., … Adesnik, H. (2018). Precise multimodal optical control of neural ensemble activity. Nature Neuroscience, 21(6), 881–893. https://doi.org/10.1038/s41593-018-0139-8
Markram, H. (2006). The Blue Brain Project. Nature Reviews Neuroscience, 7, 153. Retrieved from http://dx.doi.org/10.1038/nrn1848
Markram, H., Muller, E., Ramaswamy, S., Reimann, M. W., Abdellah, M., Sanchez, C. A., … Schürmann, F. (2015). Reconstruction and Simulation of Neocortical Microcircuitry. Cell, 163(2), 456–492. https://doi.org/10.1016/j.cell.2015.09.029
Marx, V. (2013). Neuroscience waves to the crowd. Nature Methods, 10, 1069. Retrieved from http://dx.doi.org/10.1038/nmeth.2695
Mizutani, R., Saiga, R., Takeuchi, A., Uesugi, K., & Suzuki, Y. (2013). Three-dimensional network of Drosophila brain hemisphere. Journal of Structural Biology, 184(2), 271–279. https://doi.org/https://doi.org/10.1016/j.jsb.2013.08.012
Murakami, T. C., Mano, T., Saikawa, S., Horiguchi, S. A., Shigeta, D., Baba, K., … Ueda, H. R. (2018). A three-dimensional single-cell-resolution whole-brain atlas using CUBIC-X expansion microscopy and tissue clearing. Nature Neuroscience, 21(4), 625–637. https://doi.org/10.1038/s41593-018-0109-1
Nemrodov, D., Niemeier, M., Patel, A., & Nestor, A. (2018). The Neural Dynamics of Facial Identity Processing: Insights from EEG-Based Pattern Analysis and Image Reconstruction. Eneuro, 5(1), ENEURO.0358-17.2018. https://doi.org/10.1523/ENEURO.0358-17.2018
Nishiyama, J., Mikuni, T., & Yasuda, R. (2017). Virus-Mediated Genome Editing via Homology-Directed Repair in Mitotic and Postmitotic Cells in Mammalian Brain. Neuron, 96(4), 755–768.e5. https://doi.org/10.1016/j.neuron.2017.10.004
Oizumi, M., Albantakis, L., & Tononi, G. (2014). From the Phenomenology to the Mechanisms of Consciousness: Integrated Information Theory 3.0. PLOS Computational Biology, 10(5), e1003588. Retrieved from https://doi.org/10.1371/journal.pcbi.1003588
Okano, H., Miyawaki, A., & Kasai, K. (2015). Brain/MINDS: brain-mapping project in Japan. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 370(1668). https://doi.org/10.1098/rstb.2014.0310
Peng, H., Hawrylycz, M., Roskams, J., Hill, S., Spruston, N., Meijering, E., & Ascoli, G. A. (2015). BigNeuron: Large-Scale 3D Neuron Reconstruction from Optical Microscopy Images. Neuron, 87(2), 252–256. https://doi.org/10.1016/j.neuron.2015.06.036
Peng, H., Zhou, Z., Meijering, E., Zhao, T., Ascoli, G. A., & Hawrylycz, M. (2017). Automatic tracing of ultra-volumes of neuronal images. Nature Methods, 14, 332. Retrieved from https://doi.org/10.1038/nmeth.4233
Poo, M., Du, J., Ip, N. Y., Xiong, Z.-Q., Xu, B., & Tan, T. (2016). China Brain Project: Basic Neuroscience, Brain Diseases, and Brain-Inspired Computing. Neuron, 92(3), 591–596. https://doi.org/10.1016/j.neuron.2016.10.050
Regalado, A. (2017). The Entrepreneur with the $100 Million Plan to Link Brains to Computers. MIT Technology Review. Retrieved from https://www.technologyreview.com/s/603771/the-entrepreneur-with-the-100-million-plan-to-link-brains-to-computers/
Reimann, M. W., Nolte, M., Scolamiero, M., Turner, K., Perin, R., Chindemi, G., … Markram, H. (2017). Cliques of Neurons Bound into Cavities Provide a Missing Link between Structure and Function. Frontiers in Computational Neuroscience. Retrieved from https://www.frontiersin.org/article/10.3389/fncom.2017.00048
Seo, D., Neely, R. M., Shen, K., Singhal, U., Alon, E., Rabaey, J. M., … Maharbiz, M. M. (2016). Wireless Recording in the Peripheral Nervous System with Ultrasonic Neural Dust. Neuron, 91(3), 529–539. https://doi.org/10.1016/j.neuron.2016.06.034
Song, D., She, X., Hampson, R. E., Deadwyler, S. A., & Berger, T. W. (2017). Multi-resolution multi-trial sparse classification model for decoding visual memories from hippocampal spikes in human. In 2017 39th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC) (pp. 1046–1049). https://doi.org/10.1109/EMBC.2017.8037006
Stocco, A., Prat, C. S., Losey, D. M., Cronin, J. A., Wu, J., Abernethy, J. A., & Rao, R. P. N. (2015). Playing 20 Questions with the Mind: Collaborative Problem Solving by Humans Using a Brain-to-Brain Interface. PLOS ONE, 10(9), e0137303. Retrieved from https://doi.org/10.1371/journal.pone.0137303
Suk, H.-J., van Welie, I., Kodandaramaiah, S. B., Allen, B., Forest, C. R., & Boyden, E. S. (2017). Closed-Loop Real-Time Imaging Enables Fully Automated Cell-Targeted Patch-Clamp Neural Recording In Vivo. Neuron, 95(5), 1037–1047.e11. https://doi.org/https://doi.org/10.1016/j.neuron.2017.08.011
Szigeti, B., Gleeson, P., Vella, M., Khayrulin, S., Palyanov, A., Hokanson, J., … Larson, S. (2014). OpenWorm: an open-science approach to modeling Caenorhabditis elegans. Frontiers in Computational Neuroscience. Retrieved from https://www.frontiersin.org/article/10.3389/fncom.2014.00137
Tasic, B., Yao, Z., Graybuck, L. T., Smith, K. A., Nguyen, T. N., Bertagnolli, D., … Zeng, H. (2018). Shared and distinct transcriptomic cell types across neocortical areas. Nature, 563(7729), 72–78. https://doi.org/10.1038/s41586-018-0654-5
Zheng, Z., Lauritzen, J. S., Perlman, E., Robinson, C. G., Nichols, M., Milkie, D., … Bock, D. D. (2018). A Complete Electron Microscopy Volume of the Brain of Adult Drosophila melanogaster. Cell, 174(3), 730–743.e22. https://doi.org/10.1016/j.cell.2018.06.019