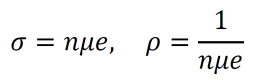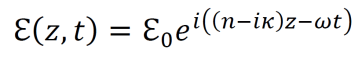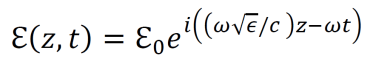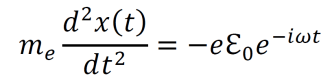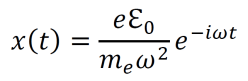Modern epistemology has generated a myriad of debatable conclusions from assumptions about knowledge and justification. For instance, René Descartes used his epistemological propositions to develop arguments for substance dualism, a philosophy which classifies the mind and the material as distinct substances (Descartes, Cottingham, & Williams, 1996). However, the possibility of an immaterial substance grows less and less likely with increased understanding of neuroscience and physics (Barrett, 2014; Oizumi, Albantakis, & Tononi, 2014). Given the almost certainly causal relationship between physical influences on the brain and cognitive changes, substance dualism seems highly unlikely at best. In addition, the existence of an immaterial substance may be inherently contradictory since any immaterial substance capable of interacting with the material world would gain materiality by virtue of its communication with the material (Poland, 1994). John Locke developed an epistemology in which he suggested that innate Ideas do not exist (Locke, 1836). But contemporary scientific data shows that many organisms do exhibit instinctual behavior patterns from birth, contradicting Locke’s position (Batki, Baron-Cohen, Wheelwright, Connellan, & Ahluwalia, 2000; Manoli, Meissner, & Baker, 2006; Schoolland, 1942). I propose that physical panpsychism may entail a form of revised empiricism, circumvent the issues associated with modern epistemology, and provide a superior framework for understanding the basis of knowledge and justification.
Panpsychism is rekindling among contemporary thinkers as an explanation for consciousness (Strawson, 2006). Integrated information theory or IIT (Oizumi et al., 2014) is a mathematical formulation which seeks to quantify consciousness using the information arising from dynamical systems. Galen Strawson has argued that IIT implies panpsychism since all physical structures contain some amount of information (Strawson, 2006). Furthermore, Adam Barrett has proposed modifications to IIT which may help account for fundamental physical interpretations of the universe like quantum field theory (Barrett, 2014). Panpsychic descriptions of reality are reentering philosophical and scientific discourse as new data are acquired and new theoretical interpretations develop.
But many still view the idea that inanimate objects may possess primitive qualia as ludicrous. To counter this presumption, consider a fragment of quartz resting on a ridge. As the sun rises, photons excite the atoms on the crystal’s surface, causing thermal oscillations to propagate into the quartz. This thermal diffusion is modulated by crystallographic defects, causing a heterogeneous distribution of heat inside the rock. As dusk falls, the quartz fragment begins to cool, emitting heat at varying rates across the surface. The particular rates are influenced directly by this quartz specimen’s pattern of internal defects. Next, consider a mouse, also located on the ridge. As the sun rises, photons excite the retinaldehyde molecules in the mouse’s eyes, triggering signal transduction via electrochemical systems. This signal moves into the mouse’s brain, where it propagates through a series of neural pathways, causing a heterogeneous distribution of neural activity. Soon, the signal’s interaction with preexisting brain structures is translated into a motor action; the mouse blinks and looks away from the bright illumination. The particular motor response is modulated by the structural organization of this mouse’s brain at the given time. The quartz and the mouse both receive sensory inputs, process them according to internal properties, and then give motor outputs. Although the rock’s “brain” is much more disorganized and chaotic than the mouse’s brain, it operates by the same basic principles and could plausibly experience a primitive form of consciousness. As such, the possibility of panpsychism cannot be readily dismissed as absurd or metaphysical.
Another prominent objection to panpsychism arises from brain processes which occur in a subconscious fashion. For instance, activity in the primary visual cortex (V1) does not correlate with conscious visual experience except for a few special cases (Boehler, Schoenfeld, Heinze, & Hopf, 2008; Boyer, Harrison, & Ro, 2005; Crick & Koch, 1995). However, the presence of subconscious neural events does not necessarily indicate that the said events are subconscious from the viewpoint of their associated anatomies. Instead, anatomical structures like V1 may experience their own independent qualia. The full informational content of their perceptions may not be transmitted or translated into the brain areas like the prefrontal cortex (PFC) which are often identified with a patient’s sense of self (Mitchell, Banaji, & Macrae, 2005). Of course, some data does transfer into higher brain regions to facilitate processes like vision, but the information undergoes an extensive series of modifications before arriving at the PFC and other regions associated with conscious processing. For this reason, the “unconscious anatomies” objection is insufficient to invalidate panpsychism.
An alternative formulation of empiricism may hold in a panpsychic universe. If all matter contains at least some primitive level of consciousness, then all channels of information transfer might be regarded as sensory. To understand this, consider the lateral geniculate nucleus (LGN), a neural structure which behaves as an intermediary point between the retina and V1 (Tortora & Derrickson, 2013). In the context of rationalism and empiricism, some may claim that only the retina provides sensory information. However, I would argue that this represents an arbitrary and ultimately falsitical distinction. Rather, I propose that every point at which data undergoes some transformation (including spatial translation) represents its own “slice” of sensory processing. As such, the LGN itself may act as a sensory organ which receives data from afferent retinal ganglia.
Much finer-grained slices of matter may contain slices of sensory processing as well. Nanoscale distances along axons contribute to the propagation of action potentials via the movement of ions. Whenever information undergoes transfer over even a miniscule distance, it acts as a sensory input into the adjacent spatial structure. In this way, the entire cosmos can be thought of as a series of interlinked sensory organs. Depending on whether physics exhibits fundamentally discrete or continuous character, there may or may not be a way of partitioning individual sensory quanta into distinct structures. It should be noted that the description of physics as sensory might be a slight misrepresentation since the term “sensory” carries connotations related to more traditional physiological concepts of sensory organs. Although every data-transfer event can be thought of as a sensory input, describing such events as a form of information processing is equally valid. Nonetheless, for the purposes of dissecting rationalism and empiricism, it is useful to describe the universe as sensory. If all data lives in a sensory field, then all knowledge and justification must be sensory. As such empiricism holds, but possesses radically different properties compared to the versions espoused by most modern empiricists.
Many distinct subsets of the universe experience intercommunication, though the level of said intercommunication is often limited. To visualize this, consider a hypothetical pair of humans named Nathaly and Ernesto. Nathaly does not possess the ability to read Ernesto’s mind. However, Ernesto may explain the content of his mind verbally. Despite this explanation representing an incomplete dataset which undergoes heavy transformation as he maps the information into sound waves and Nathaly maps the sound waves back into patterns of neural firing, some of Ernesto’s mental content is roughly reconstructed in Nathaly’s brain. I suggest that this type of interaction lies at the heart of knowledge and justification. Informational connectivity provides an empirical link by which subsystems of reality create representations of other subsystems and gain knowledge of the universe.
On a larger scale, knowledge and justification may permeate the cosmos, but in a mostly stochastic and primitive fashion. Any given subset of reality may possess some knowledge and justification about itself and about other subsets of reality. However, most structures likely exhibit extremely limited understanding of external data. For example, consider a cloud of dust particles drifting in outer space. The cloud may possess a very primitive level of consciousness. I speculate that such an entity’s experience may resemble the static white noise which sometimes appears on television screens. This cloud may acquire knowledge and justification by thermal excitation from incoming starlight. The thermal excitation represents a direct causal influence from the stars onto the dust cloud. As a result, the cloud’s state undergoes an alteration with a direct relationship to the states of the stars. Of course, the cloud’s cognitive simulation of the distant stars is vastly different from the kind of cognitive simulation a human would experience, though some new qualia likely occurs in the cloud relative to its unexcited state. Nonetheless, the cloud possesses a form of knowledge. To extend this concept, the cloud only has a very specific form of justification for this knowledge. The cloud gains empirical justification that it experiences a state change, but does not possess the level of cognitive machinery necessary to develop a veridical epistemic model of the distant stars. Subsystems within the universe continuously generate empirical models of other subsystems, but the epistemic precision of said models varies depending on the cognitive construction of the given subsystem.
More intelligent structures may possess superior epistemic abilities. Here, I specifically refer to the type of intelligence which allows a system to create accurate representative models of other structures. As an illustrative example, imagine that a group of engineers builds an “empathy machine.” The empathy machine moves about and gathers cellular-resolution scans of people’s functional brain activity. After a scan, the empathy machine simulates the person’s brain with exquisite detail, creating a near-perfect replica of that person’s experiences. In this way, the empathy machine can acquire veridical knowledge of other subsystems. Its knowledge and justification is empirical, the product of sensory experience, but possesses far more accuracy than the dust cloud’s knowledge from the scenario described previously. Recall that sensory experience extends beyond sensory organs and includes every data-transfer event within an intelligent system. Note that the veridicality of knowledge and justification may or may not be practically important. I emphasize veridicality here in order to provide a descriptive philosophical classification scheme rather than to make claims about epistemic virtue. The empathy machine shows that appropriately organized systems may acquire fairly complex and veridical information about each other.
Although the majority of subsystems within the universe only propagate distorted versions of their own experiential patterns into neighboring subsystems, the cosmos may have the potential for vastly greater connectivity. In a possible future scenario, the universe may reach a point at which it is saturated with engineered intelligence (Kurzweil, 2005). If all matter is incorporated into programmable computronium structures, intercommunication processes may allow any subset of reality to accurately simulate any other subset of the reality. Distant subsets may communicate with relative efficiency by altering the spacetime metric since changing distance itself does not violate the lightspeed barrier (Alcubierre, 1994). Even in this scenario, simultaneous sensory knowledge of all states in the cosmos may present a challenge since a limited amount of computational resources can be packed into given spatial bounds (Bremermann, 1967) and so a limited amount of knowledge could undergo simulation at any given spatiotemporal location. However, a possibility remains for the universe to behave as an enormous brain, partitioning its attentional mechanisms to different subsets of the total knowledge and justification as the need arises. Furthermore, an entity on this scale would likely have abilities which current humans cannot conceptualize, so the constraints involved might be very different than those described here. This extreme situation demonstrates that the most crucial limit to empirical knowledge and justification arises from informational connectivity and the capacity for state emulation.
Using panpsychism as an explanation for the existence of consciousness in a physical universe, an alternative form of empiricism emerges. Sensory knowledge can be reframed as events by which information undergoes transformation. For this reason, any cognitive process might be considered empirical, regardless of whether said process is actually associated with traditional sensory organs. Distinct subsystems within the cosmos gain knowledge and justification by receiving data from each other and undergoing state alterations which modify their qualia. Veridicality is achieved when the state alterations emulate the states of the structure which induced the change. If the entire universe was engineered to maximize informational connectivity, it may acquire a form of omniscient or nigh-omniscient understanding in which any knowledge can be accessed and understood as needed. Panpsychic epistemology provides a new perspective on knowledge and justification which may facilitate the convergence of scientific and philosophical approaches to epistemological questions.
References
Alcubierre, M. (1994). The warp drive: hyper-fast travel within general relativity. Classical and Quantum Gravity, 11(5), L73. Retrieved from http://stacks.iop.org/0264-9381/11/i=5/a=001
Barrett, A. (2014). An integration of integrated information theory with fundamental physics. Frontiers in Psychology. Retrieved from https://www.frontiersin.org/article/10.3389/fpsyg.2014.00063
Batki, A., Baron-Cohen, S., Wheelwright, S., Connellan, J., & Ahluwalia, J. (2000). Is there an innate gaze module? Evidence from human neonates. Infant Behavior and Development, 23(2), 223–229. http://doi.org/https://doi.org/10.1016/S0163-6383(01)00037-6
Boehler, C. N., Schoenfeld, M. A., Heinze, H.-J., & Hopf, J.-M. (2008). Rapid recurrent processing gates awareness in primary visual cortex. Proceedings of the National Academy of Sciences, 105(25), 8742 LP-8747. Retrieved from http://www.pnas.org/content/105/25/8742.abstract
Boyer, J. L., Harrison, S., & Ro, T. (2005). Unconscious processing of orientation and color without primary visual cortex. Proceedings of the National Academy of Sciences of the United States of America, 102(46), 16875 LP-16879. Retrieved from http://www.pnas.org/content/102/46/16875.abstract
Bremermann, H. J. (1967). Quantum noise and information. In Proceedings of the Fifth Berkeley Symposium on Mathematical Statistics and Probability, Volume 4: Biology and Problems of Health (pp. 15–20). Berkeley, Calif.: University of California Press. Retrieved from https://projecteuclid.org/euclid.bsmsp/1200513783
Crick, F., & Koch, C. (1995). Are we aware of neural activity in primary visual cortex? Nature, 375(6527), 121–123.
Descartes, R., Cottingham, J., & Williams, B. (1996). Descartes: Meditations on First Philosophy: With Selections from the Objections and Replies. Cambridge University Press.
Kurzweil, R. (2005). The Singularity Is Near: When Humans Transcend Biology. New York: Viking.
Locke, J. (1836). An essay concerning human understanding. T. Tegg and Son.
Manoli, D. S., Meissner, G. W., & Baker, B. S. (2006). Blueprints for behavior: genetic specification of neural circuitry for innate behaviors. Trends in Neurosciences, 29(8), 444–451. http://doi.org/https://doi.org/10.1016/j.tins.2006.06.006
Mitchell, J. P., Banaji, M. R., & Macrae, C. N. (2005). The Link between Social Cognition and Self-referential Thought in the Medial Prefrontal Cortex. Journal of Cognitive Neuroscience, 17(8), 1306–1315. http://doi.org/10.1162/0898929055002418
Oizumi, M., Albantakis, L., & Tononi, G. (2014). From the Phenomenology to the Mechanisms of Consciousness: Integrated Information Theory 3.0. PLOS Computational Biology, 10(5), e1003588. Retrieved from https://doi.org/10.1371/journal.pcbi.1003588
Poland, J. (1994). Physicalism, the Philosophical Foundations (Vol. 57). Oxford University Press.
Schoolland, J. B. (1942). Are there any innate behavior tendencies? Genetic Psychology Monographs, 25, 219–289.
Strawson, G. (2006). Realistic monism: why physicalism entails panpsychism.
Tortora, G. J., & Derrickson, B. H. (2013). Principles of Anatomy and Physiology, 14th Edition: 14th Edition. Wiley Global Education.









")