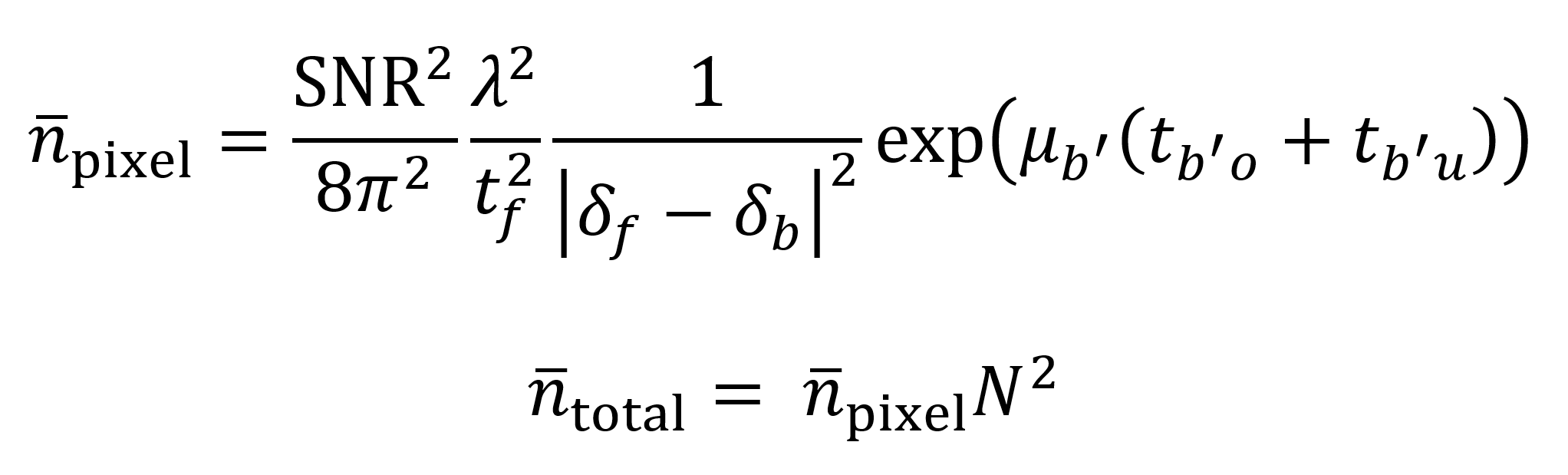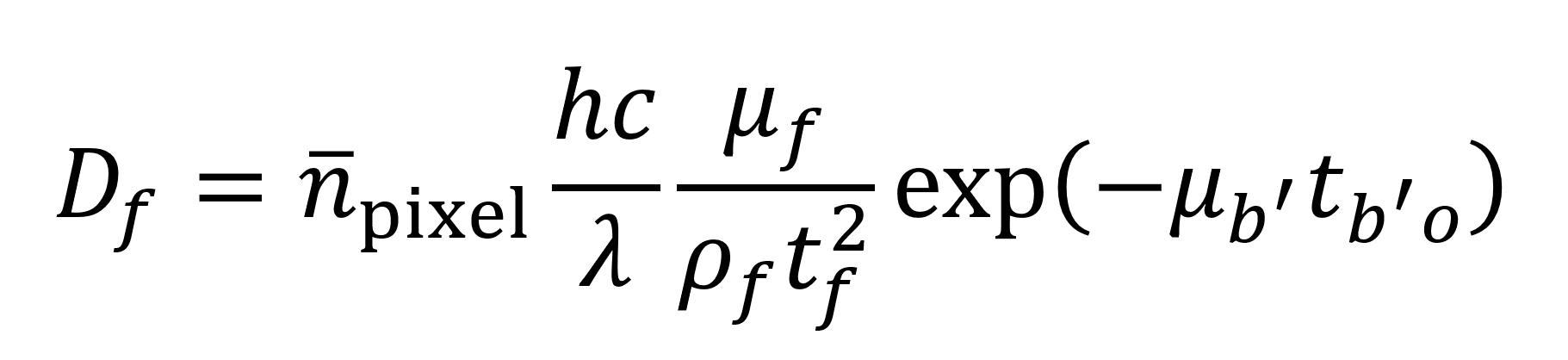During my pursuits, I’ve come across an increasing number of exciting nontraditional routes for funding scientific research. The efforts of Adam Marblestone and Benjamin Reinhardt have been particularly instrumental in stimulating this ecosystem, but many other great people have contributed as well. These new funding routes are a welcome relief since many of the most innovative and far-reaching projects are not especially suited for receiving governmental NIH, NSF, etc. funding. If you would like to find a more comprehensive list of such alternative funding sources, you should check out https://arbesman.net/overedge/. My own list (below) consists of funding sources that stand out to me as particularly promising. I hope you find this useful and feel free to reach out if you have any questions!
Amaranthe Foundation https://amaranth.foundation/bottlenecks-of-aging “We outline initiatives which, if executed, could meaningfully accelerate the advancement of aging science and other life-extending technologies. The resulting document is a philanthropic menu, for which Amaranth is seeking both talent to execute on and co-funders. If you are a founder, researcher, or philanthropist interested in executing or co-sponsoring one or several of the projects or proposals below, please reach out to us”.
Arc Institute https://arcinstitute.org/ “Headquartered in Palo Alto, California, Arc is a nonprofit research organization founded on the belief that many important scientific programs can be enabled by new organizational models. Arc operates in partnership with Stanford University, UCSF, and UC Berkeley. Arc gives scientists no-strings-attached, multi-year funding, so that they don’t have to apply for external grants and invests in the rapid development of experimental and computational technological tools. As individuals, Arc researchers collaborate across diverse disciplines to study complex diseases, including cancer, neurodegeneration, and immune dysfunction. As an organization, Arc strives to enable ambitious, long-term research agendas. Arc’s mission is to accelerate scientific progress, understand the root causes of disease, and narrow the gap between discoveries and impact on patients.”
ARIA (Advanced Research + Invention Agency) https://www.aria.org.uk/ “ARIA empowers scientists to pursue breakthroughs at the edge of the possible. We have a big mission, and getting there won’t be easy. That’s why we’re building a new kind of research agency that does things differently. Created by an Act of Parliament, and sponsored by the Department for Science, Innovation, and Technology, ARIA will fund projects across the full spectrum of R&D disciplines, approaches, and institutions. ARIA’s programmes and projects are directed by our Programme Directors, scientific and technical leaders with deep expertise and a focused, creative vision for how technology can enable a better future. While Programme Directors are tasked with deeply exploring a topic and designing funding opportunities, they won’t get to breakthroughs alone. To maximise their chance at success, they’ll develop their thinking through direct calls for feedback, source projects through open solicitations, and have their programmes reviewed by experts against a clear set of evaluation criteria.”
Astera Institute https://astera.org/“We empower visionary, high-leverage science and technology projects with the capacity to create transformative progress for human civilization. We target programs in Artificial General Intelligence, Science, and Climate that currently lack a natural home in the existing innovation ecosystem.”
Brains https://spec.tech/brains “Brains is a training program to provide the skills and opportunities to translate ambitious research visions that aren’t a good fit for a company but are too big for a single academic lab into impact. These visions could be anything from upending the way we make carbon-based products to how we understand the brain or build air- breathing fusion engines. Think YCombinator for coordinated research programs.”
Convergent Research https://www.convergentresearch.org/ “New types of organization are needed to accelerate scientific progress. Academic research groups and startup companies are essential to science and technology development. But there are some projects they just aren’t suited for. A university astronomy lab couldn’t have launched the Hubble Space Telescope on its own, nor would a venture-backed startup have built the Large Hadron Collider at CERN. Hubble and CERN illustrate a common pattern in science: a need for projects that are bigger than an academic lab can undertake, more coordinated than a loose consortium or themed department, and not directly profitable enough to be a venture-backed startup or industrial R&D project. Focused Research Organizations (FROs) are a new type of scientific institution designed to fill this gap.”
Emergent Ventures Grant/Fellowship https://www.mercatus.org/emergent-ventures “We want to jumpstart high-reward ideas—moonshots in many cases—that advance prosperity, opportunity, liberty, and well-being. We welcome the unusual and the unorthodox. Our goal is positive social change, but we do not mind if you make a profit from your project. (Indeed, a quick path to revenue self-sufficiency is a feature not a bug!) Projects will either be fellowships or grants: fellowships involve time in residence at the Mercatus Center in Northern Virginia; grants are one-time or slightly staggered payments to support a project. We encourage you to think big, but we also will consider very small grants or short fellowships if they might change the trajectory of your life. We encourage applications from all ages and all parts of the world.”
Flux Capacitor https://1517.substack.com/p/the-flux-capacitor-time-funds-and “Flux Capacitor is a 3-month break away from academia to pursue out-there ideas to build into a startup OR moonshot science… You want to hit pause on the academic rat race and spend 3 months on first-principles exploration of either applied, practical problems that can be commercialized within a VC-funded startup in the near-medium term (5 years) or moonshot fundamental science. To help you do this, we’ll give you up to $100k in funding.”
Foresight AI Safety Grant Program https://foresight.org/ai-safety/ “This grant program seeks to support projects working to make progress on three areas we consider underexplored when it comes to AI Safety… 1. Neurotechnology, Whole Brain Emulation and lo-fi Uploading for AI safety; 2. Security, Cryptography, and Auxiliary Approaches for Infosec and AI Security; and 3. Safe and Beneficial Multipolar AI Scenarios… Projects will be evaluated by a mix of Foresight staff and external advisors. We aim to focus on projects that have a chance of being successful within short AI timelines. Rather than funding many projects with the potential of making a small difference in the long-run, we may be more inclined to fund projects that are high-risk high-reward, in the sense that they are more speculative but would make a big difference if successful. Generally, we are interested in proposals for scoping/mapping opportunities in this area, especially from a differential technology development perspective.”
Hypothesis Fund https://www.hypothesisfund.org/ “The Hypothesis Fund advances scientific knowledge by supporting early stage, innovative research that increases our adaptability against systemic risks to the health of people and the planet. We make seed grants to fund research projects at their earliest stages, typically before there is any preliminary data. Our funding is intended to be catalytic — a fast path to enable a scientist to ‘turn over the card’ and see what’s there. And we focus on bold new ideas in basic research, not continuations of existing research. The Hypothesis Fund approach is different. We empower a world-class and diverse network of scientist Scouts to identify the high-risk, high-reward ideas at the edge of the network that would otherwise be un-pursued or underfunded.”
Long-Term Future Fund https://funds.effectivealtruism.org/funds/far-future “The Long-Term Future Fund aims to positively influence the long-term trajectory of civilization by making grants that address global catastrophic risks, especially potential risks from advanced artificial intelligence and pandemics. In addition, we seek to promote, implement, and advocate for longtermist ideas, and to otherwise increase the likelihood that future generations will flourish.”
OpenResearch https://www.openresearchlab.org/ “OpenResearch is a nonprofit research lab. We fund work that requires a very long time horizon, seeks to answer open-ended questions, or develops technology that shouldn’t be owned by any one company.”
Renaissance Philanthropy https://renaissancephilanthropy.org/ “Renaissance Philanthropy’s mission is to fuel a 21st century renaissance by increasing the ambition of philanthropists, scientists, and innovators. We do this by advising philanthropists, surfacing breakthrough ideas, and incubating ambitious initiatives. Our aim is to activate a virtuous loop of increasing ambition and impact between philanthropists and innovators: by identifying frontier experts both in science and in new ways of solving problems; by tapping into the growing number of emerging philanthropists; and by building multi-sector initiatives that can harness the power of philanthropy, markets, and governments.”
Survival and Flourishing Fund https://survivalandflourishing.fund/speculation-grants.html “SFF Speculation Grants are expedited grants organized by SFF outside of our biannual grant-recommendation process (the S-process). “Speculation Grantors” are volunteers with budgets to make these grants. Each Speculation Grantor’s budget grows or increases with the settlement of budget adjustments that we call “impact futures” (explained further below). Currently, we have a total of ~20 Speculation Grantors, with a combined budget of approximately $10MM (up from $4MM initially). Our process and software infrastructure for funding these grants were co-designed by Andrew Critch and Oliver Habryka.”
1517 Fund https://www.1517fund.com/ “1517 is a venture capital fund and community supporting college dropouts, renegade students, and deep tech scientists with investment at the earliest stages of their companies. Founded by the cofounders of the Thiel Fellowship, it supports founders across software, hardware, and deep tech verticals and also provides a community to hackers, makers, and scientists from across the world.”
I’m compiling this list to agglomerate just a few of the resources that I find useful for learning about how the world works, developing new skills, expanding my repertoire of ways of looking at global challenges, and planning how to contribute towards creating the future. Since many of the links here might change over time, please comment if you find any that do not work so that I can look for suitable replacement links. Also, feel free to let me know if you know of good resources not listed here! I’m always excited to expand my learning.
Bottlenecks of Aging is a funding effort by the Amaranth Foundation which has selected a set of 12 limiting facets that have hindered progress towards extending the healthy human lifespan and is seeking proposals for projects which address these priorities.
Convergent Research is a group devoted to spinning off Focused Research Organizations (FROs). An FRO is typically awarded around $30M to pursue a midsize research goal over the course of 5 years. FROs focus on goals that are too ambitious for academia but do not have a near-term commercial incentive that would make them work well as startup companies. Convergent Research was co-founded by Adam Marblestone.
Emergent Ventures Grant/Fellowship is a program run by philanthropist Tyler Cowen which offers funding to applicants pursuing exciting projects. Anyone over the age of 13 can apply and the application process is straightforward. The Fellowship option additionally gives awardees the opportunity to receive mentorship through time in residence at the Mercatus Center in Northern Virginia.
F99/K00 Transition to Aging Research for Predoctoral Students is an NIH grant to which graduate students can apply in the last few years of their PhD training. It provides funding for the final 1-2 years of the PhD as well as for up to 4 years of postdoctoral research.
NIH’s grants homepage is a website by the American National Institutes of Health which provides information about their grant programs.
Speculative Technologies is a nonprofit founded by Benjamin Reinhardt which funds “systems research” that does not fit easily into academia or industry. This means research that is necessary to create multiple interacting technologies that enable new capabilities to serve society. Unlike the FROs of Convergent Research, projects funded by Speculative Technologies are parallelized across multiple institutions and are not necessarily defined by a single approach to start.
The Foresight Institute’s AI Safety grant program provides funding to project proposals that address certain underexplored areas of AI safety including whole-brain emulation and neurotechnology, multipolar AI scenarios, and information security. It is sponsored by the Foresight Institute.
The Overedge Catalog is a webpage listing a variety of non-traditional funding opportunities for science and technology research.
Understanding science funding in tech is a blog page by Nadia Asparouhova which discusses the history, approaches, and future of efforts to create new institutions for scientific funding which properly encourage innovation and circumvent the weight of bureaucracy present in existing federal funding sources.
1517 Fund is a nontraditional venture capital firm that backs “dropouts, renegade students & sci-fi scientists at the earliest stages of their companies”, particularly in deep tech fields. It features a variety of options for funding and reaching out to them to discuss specifics is encouraged.
Entrepreneurship and Organization Management
Erika’s quick-start guide to research nonprofits is a blog page by Erika Alden Debenedictis that gives information on how to raise funding from philanthropists, manage money, manage IP and spinouts, hire people, get lab space, and more.
Notes on Starting a Scientific Organization is a blog page by Sabrina Singh which provides information on how to start and manage new scientific organizations. It is particularly useful for learning about the bookkeeping and staffing aspects of management.
So you want to start a biotech company is an article published in Nature Biotechnology that goes over some key concepts important for entrepreneurship in the biosciences.
Trailblazer List is a website that describes numerous companies working in areas (e.g. neurotechnology, sustainability, computing, fabrication, and many more) that have the potential to radically impact the future for the better.
Venture Capital for Bio 101 is a blog page by Celine Halioua which goes over the process of obtaining VC funding for biotechnology startups as well as some strategies to use and pitfalls to avoid.
History, Science, Society, and the Future
BigThink The Progress Issue is a special issue of the science and society news website organization known as BigThink and its similar partner site FreeThink. This issue is devoted to articles that use data to support rationally optimistic approaches to creating the future and that emphasize the importance of leveraging data-driven hope to overcome the predominant cultural zeitgeist of pessimism.
Caspian Report is an educational YouTube channel on geopolitics. In my opinion, it is a reputable source as YouTube channels go and presents information in a minimally biased fashion.
Future of Humanity Institute is a multidisciplinary research organization at the University of Oxford which combines philosophy, mathematics, and social science techniques to study existential risks, long-term future scenarios, and how humanity might best move forward. It was founded by Professor Nick Bostrom and employs Toby Ord (co-founder of the Effective Altruism movement) and Professor Anders Sandberg as senior researchers.
Future of Life Institute is a nonprofit organization with a mission of “steering transformative technology towards benefiting life and away from extreme large-scale risks”. It works through policy development and advocacy, education and outreach, research and grantmaking, institution building, and coordinating conferences. It focuses primarily on existential risks posed by artificial intelligence (esp. autonomous weapons), biotechnology, and nuclear weapons.
Matt Bell is a blog that has a number of excellent articles on science, futurism, and other topics. It has a particularly compelling post on Embracing the Cosmic Endowment.
Our World in Data is a website that collects reputable data about the state of the world, organizes it into various categories, and provides useful ways of visualizing said data.
Science X is a consortium of science news websites including Phys.org, Medical Xpress, and Tech Xplore. These sites post massive numbers of articles summarizing scientific papers and other types of advances in an accessible fashion.
The Library of Existential Hope is a website associated with the Foresight Institute which compiles resources on existential hope and existential risk management as well as on technology areas with the potential to shape the future.
Computing
Digital Fundamentals is a textbook by Thomas Floyd that discusses the hardware architecture and mechanisms of digital computers. In my view, it has excellent visuals and explains concepts very clearly.
HPCWire is a news organization that writes articles about cutting-edge developments in the high-performance computing industry.
Reducible is a YouTube channel that uses animations and excellent teaching to explain concepts in computer science such as algorithms, image processing, and applied graph theory.
Synthetic Biology and Biotechnology
Addgene is a company that distributes and sells plasmids (at reasonable prices) made by laboratories around the world. It represents a central repository for a wide variety of useful plasmids that can aid biotechnology research.
Addgene’s Viral Plasmids and Resources page provides useful articles that describe the basics of lentivirus, adeno-associated virus (AAV), adenovirus, and γ-retrovirus as well as links to some other resources.
AlphaFold Protein Structure Database is a searchable catalog of predicted protein structures along with information about the accuracy of each prediction. Most known proteins are represented in the database. This catalog was created using the spectacularly successful AI protein prediction software AlphaFold. Although not all of the predictions are perfect and many include high uncertainty within intrinsically disordered regions, the data most often agree strongly with experimental validations.
GEN Genetic Engineering & Biotechnology News is an excellent magazine that covers cutting-edge topics in the biotechnology industry. It is aimed at a scientific audience but might be somewhat accessible to others as well.
GROMACS documentation is an online manual describing how to use the GROMACS molecular dynamics simulation software package.
J. Craig Venter Institute is a nonprofit research organization that focuses on synthetic biology, building minimal cells, and developing genomics technologies. It is led by the National Medal of Science recipient J. Craig Venter, one of the main contributors to the success of human genome sequencing.
National Center for Biotechnology Information (NCBI) is a collection of bioinformatics databases and tools sponsored by the NIH. The website hosts massive libraries of biological data such as DNA sequences, protein sequences, and many more. It also includes BLAST, a popular search tool that facilitates comparison of the evolutionary similarities among different biomolecular sequences.
Oxman is the website of a company founded by the transdisciplinary designer-biologist Professor Neri Oxman of the MIT Media Lab. This website details some of the remarkable work by Neri Oxman and her team on devising ways of redesigning the built world to more seamlessly interact with the rest of the biosphere. They utilize architecture, design, art, philosophy, computer science, synthetic biology, and more to develop innovative new materials and systems.
RCSB Protein Data Bank (PDB) is a website that catalogs experimentally determined structures of proteins and protein complexes. It is user friendly and includes a wide range of features for analyzing its protein structures.
SynBioBeta is an “innovation network” for bioengineers, entrepreneurs, and investors in the synthetic biology industry, particularly within the area of sustainability. It hosts a yearly conference called The Global Synthetic Biology Conference. In addition, its website includes an industry news section that posts synthetic biology news articles.
Bioprocess International is an industry news website with articles that discuss biomanufacturing, biological purification methods, analytical techniques, and similar topics.
Insights On Successful Gene Therapy Manufacturing And Commercialization is a booklet compiled by industry experts on the logistical and technical challenges surrounding large-scale manufacturing of viral vectors for clinical gene therapy as well as on emerging solutions to these difficulties.
Neuroscience and Neurotechnology
Allen Brain Map is a website hosting a treasure trove of many different neurobiology databases (such as the cell types database) developed through the multifaceted investigations of the Allen Institute.
BossDB (Brain Observatory Storage Service & Database) is a publicly accessible data repository for storing and disseminating petabyte-scale neuroscience data, particularly from volume electron microscopy and x-ray microtomography.
Expansion Microscopy: Super-Resolution Imaging with Hydrogels is a review paper published in the journal Analytical Chemistry which discusses the methodologies of expansion microscopy (ExM), a powerful technique for imaging tissues. ExM works by infusing tissues with a swellable hydrogel to physically enlarge them in an isotropic fashion. This particular review paper was written by Sven Truckenbolt, one of the lead scientists at E11 Bio.
Global Highlights in Neurotechnology, Connectomics, and Brain Simulation: 2005 to 2019 is my own compilation of roughly one paragraph descriptions of scientific and organizational advances in forward-thinking neuroscience topics. At this point, it is somewhat outdated since it only covers advances up to 2019. Nonetheless, it is quite useful for reviewing the history and progress of these fields.
Human Brain Project is a large-scale European consortium that leverages a variety of computational and experimental approaches to better understand neuroscience. It grew out of Henry Markram’s Blue Brain Project at EPFL which focuses more on computational neuroscience.
McGovern Institute is an institute at MIT which focuses on neuroscience and neurotechnology. Some of the world’s top researchers such as Ed Boyden and Feng Zhang lead research groups as part of this institute.
NIH BRAIN Initiative (Brain Research Through Advancing Innovative Neurotechnologies) is an American governmental program that funds efforts towards more comprehensive understanding of neuroscience.
Preventing antisocial robots: A pathway to artificial empathy is a viewpoint journal article published in Science Robotics which proposes ways of encoding affective empathy (not just cognitive empathy) into AI to prevent machines from acting as sociopaths.
The Singularity Is Near: When Humans Transcend Biology is a foundational text in futurism that proposes arguments that humanity is on the brink of a tipping point called the Singularity where accelerating technological progress might drive profound changes in the human condition. It focuses on artificial intelligence in particular as a major force behind potential dramatic changes to our world (and even universe) in the coming decades. Though this text is thought by many to make some rather unlikely claims, it presents valuable ideas to contemplate even if some of them seem outlandish.
Longevity
Hallmarks of aging: An expanding universe is a review paper that provides an excellent overview of the biology of aging and efforts to treat the condition. This paper represents a 2023 update based on a seminal 2013 review titled The Hallmarks of Aging.
Human Ageing Genomic Resources is a website that collects searchable databases of aging-related genes, drug data, variants, and more. It aids and facilitates quantitative research on aging topics.
Nature Aging is a top scientific journal that publishes scientific papers across the aging research fields.
Oviva is a startup company developing therapeutics to improve ovarian function, prevent menopause, and extend healthspan in women. Co-founded by Daisy Robinton, Oviva aims to bring women’s health into the longevity conversation.
Protective and Enhancing Alleles is a list of known human alleles compiled by George Church’s lab that are known to contribute to various abilities such as disease resistance, longevity, strength, intelligence, etc.
Want to live to 150? The world needs more humans is an editorial originally published in the Washington Post which details compelling ethical arguments in favor of healthspan extension. Though paywalled at the Washington Post, it is available for free from the author Raiany Romanni at her website.
Mathematics
Quanta Magazine is an online magazine that publishes educational articles on topics in mathematics, computer science, physics, and biology. These articles are written in an accessible style yet emphasize rigorous mathematical approaches to questions.
3Blue1Brown is an educational YouTube channel that covers advanced mathematics in an accessible way by using beautiful animations and top-notch teaching to illustrate precise quantitative concepts which might otherwise be much more challenging to understand (e.g. 10-dimensional sphere packing, the math behind deep learning, etc.)
Understanding Topology: A Practical Introduction is a remarkably accessible textbook on topology. It provides the clearest explanations (in my view) out of all of the many topology books I’ve examined.
Hedonistic Imperative is a book-length online manifesto by the philosopher David Pearce which makes philosophical arguments in favor of abolishing all suffering by using future biotechnology. He further argues for radically enhancing positive emotions while retaining motivation through “gradients of bliss”. Within his manifesto, Pearce makes a compelling case for sustainably eliminating suffering in wild animals.
Here’s How to Teach Yourself Physics and Math is a webpage on Futurism.com that links to several different lists of textbooks which one can use to learn physics and mathematics from beginner to advanced levels.
Notes on Classical Mechanics is my own webpage with explanations of Lagrangian and Hamiltonian mechanics as well as linear oscillations and coupled linear oscillators.
Notes on Optics and Microscopy is my own webpage with some explanations of optical physics and its applications in microscopy.
Earth and Space
[C]Worthy is a seed-stage Focused Research Organization that develops methodologies for monitoring, reporting, and verifying the effects of marine carbon dioxide removal technologies. They are building software for multiscale oceanographic modeling and data integration that will quantify the efficacy and the ecological impacts of marine carbon dioxide removal. They aim to provide the validation framework necessary as part of efforts to enable gigaton-scale carbon dioxide removal by 2050.
SpaceBorn United is an organization engaged in research on ways of enabling human reproduction in outer space. They have developed a prototype “space embryo incubator” and have a roadmap of space mission plans to facilitate their research on developmental biology in space. They have engaged broad expertise and partners to help address technical, medical, regulatory/legal, and ethical challenges associated with their goals.
Cover image: created using Midjourney generative AI.
Notes: males are tiny compared to females. In many species of anglerfish, mating occurs through males attaching to and then fusing with females such that their circulatory systems join together. The male provides sperm and is eventually absorbed into the female.
Notes: second largest type of shark (after the whale shark), reaches about 7.9 m in length. They feed on plankton by swimming forwards with the mouth open and filtering via gill rakers.
Notes: characterized by transparent flaps connecting the dorsal and dorsolateral tentacles in adult females. In a case of extreme sexual dimorphism, the females grow to around 2 m in size while the males are only about 2.4 cm.
Notes: at around 10 m long, they are shorter than giant squid, but they are much heavier. Colossal squid also has the largest eyes of any organism (around 30-40 cm in diameter).
Notes: depending on the type crinoid adults can either swim freely or be tethered to the sea floor by a stalk. The former are called feather stars and the latter are called sea lillies. Crinoids can also crawl using rootlike structures called cirri as legs. They consume plankton and detritus by filtering through their featherlike arms and then propelling it towards a mouth. They reproduce sexually, releasing sperm and eggs into the water. Fertilized eggs hatch into freely swimming larvae that settle on the sea floor and transition into a stalked juvenile state before eventually breaking away (in the case of feather stars) as adults to swim freely once more.
Notes: cuttlefish are among the most intelligent invertebrates. They can rapidly change color using their chromatophore cells as a mode of communication and camouflage as well as to warn off predators (called a deimatic display).
Notes: garden eels are distinguished by their behavior of living in burrows on the sea floor and poking their heads out of the burrows to eat prey and sliding back into the burrows to avoid predators. Colonies of them can resemble grasses, hence their name.
Notes: a type of small (up to 3 cm) sea slug that floats upside down using a gas-filled stomach to stay adrift. They feed on Portuguese man o’ war jellyfish and similar organisms. After consuming venomous nematocysts from their prey, they store the stinging cells in sacs (called cnidosacs) located in their own extremities. This concentrates the nematocysts, making the sting of Glaucus atlanticus potentially more potent even than that of the man o’ war jellyfish itself.
Notes: one of the largest types of jellyfish. Though their sizes vary widely, some can reach a bell diameters of more than 2 m and possess tentacles extending for over 30 m in length.
Notes: a category of animals that exist as flat sheets a few cells thick with a ciliated epithelium on their undersides. They use these cilia to move along the seafloor and most of them reproduce asexually by budding or fragmenting into smaller individuals (though one subtype does also reproduce sexually).
Notes: large sea stars which can reach diameters of 1 m. Sunflower sea stars are predatory and consume various prey such as sea urchins, other sea stars, clams, sea cucumbers, and more. They move at a speed of about a meter per minute using thousands of tube feet located on their undersides.
Notes: a deep sea cephalopod of about 30 cm length that lives in the ocean’s aphotic zone. Vampire squid (Vampyroteuthis infernalis) have flaps of tissue connecting their tentacles, each of which is lined by fleshy spines. It is covered in photophores that produce disorientating flashes of light to confuse predators. Rather than ink, they can eject a sticky cloud of bioluminescent mucus when highly agitated by predators.
Humongous Fungus, a specimen of Armillaria ostoyae, has claimed the title of world’s largest single organism. Though it features honey mushrooms above ground, the bulk of this creature’s mass arises from its vast subterranean mycelial network of filamentous tendrils. It has spread across more than 2,000 acres of soil and weighs over 30,000 metric tons. Yet I would contend that Humongous Fungus represents a mere microcosm of the world’s true largest organism, a creature that I will call Cyborg Earth. What is Cyborg Earth? Eastern religions have suggested that all life is fundamentally interconnected. Cyborg Earth represents an extension of this concept.
All across the globe, biological life thrives. Quintillions upon quintillions of biomolecular computations happen every second, powering all life. Mycoplasma bacteria. Communities of leafcutter ants. The Humongous Fungus. Beloved beagles. Seasonal influenza viruses. Parasitic roundworms. Families of Canadian elk. Vast blooms of cyanobacteria. Humanity. Life works because of complexity that arises from simplicity that in turn arises from whatever inscrutable quantum mechanical rules lay beneath the molecular scale.
All creatures rearrange atoms in various ways. Termites and beavers rearrange larger bunches of atoms than most organisms. As humans progressed from paleolithic to metalwork to industrialization and then to the space age, information revolution, and era of artificial intelligence, they learned to converse with the atoms around them in an ever more complex fashion. We are actors in an operatic performance, we are subroutines of evolution, we are interwoven matryoshka patterns, an epic chemistry.
Thermodynamics and memetic natural selection juggle our civilization while riding a unicycle along a path to an as-yet unknown destiny. Because of humanity, technology represents a fundamental part of the biosphere. Computers, airplanes, factories, shipping routes. All of it is natural. This does not mean it is good or bad. Simply that it comprises part of the great biological conversation. This is Cyborg Earth. The world’s largest organism is the biosphere itself, including all of the remarkable ways that its constituents have reshaped its body.
Cyborg Earth has always been a uniquely gleaming gem in the cold vastness of the cosmos. But I believe that Cyborg Earth still resides in an embryonic state. As we hurtle into the expanse of the infinite future, Cyborg Earth will grow and change. We must realize that we are all connected by the electric dance of atoms. As a constituent species within Cyborg Earth, we currently possess an enormous power and a tremendous responsibility to properly steward our world’s embryogenesis.
Cover image modified from “Confocal 3D-image of a fungal network with reproductive spores containing nuclei” by Vasilis Kokkoris, source.
Announcement: I have completed an early-stage study seeking to feasibilize expansion X-ray microscopy. You can access my preprint here on arXiv. My study demonstrated a metallic gold Unclearing method for generating contrast in expanded tissues. It should be noted that this study was only been able to resolve cell bodies and not neurites. However, there are likely ways of overcoming this obstacle as detailed in the preprint’s discussion section. While much work remains to be done, I hope that my findings will provide a foundation for future research to build upon. If you are interested in expansion X-ray microscopy, please feel free to reach out to logan (dot) phospholipid (at) gmail (dot) com.
Supplemental MATLAB code is located at the end of this page before the references
Combining synchrotron x-ray microscopy with expansion microscopy may represent a feasible approach for whole human brain connectomics at the nanoscale. Synchrotron x-ray microtomography on its own provides extremely fast imaging at high resolution, yet necessary tradeoffs between imaging throughput and resolution mean that the imaging an entire human brain with voxel sizes of less than 30 nm may still take much too long with current synchrotron technology. Fast imaging with voxel sizes of 300 nm is much more readily achievable. Furthermore, expansion microscopy isotropically enlarges tissue by infusion of a swellable hydrogel, facilitating resolution increases. The combination of x-ray microtomography and expansion microscopy (hereafter referred to as ExxRM) could thus push the effective voxel size down to the level needed for dense connectomics. However, because tissue volume and imaging time scale cubically with expansion factor, careful balance between design of the synchrotron x-ray optical setup and the degree of expansion will be needed. In particular, large high-resolution detectors will be vital. Additionally, methods to compensate for signal dilution from expansion and to stabilize expanded tissue under brilliant x-rays will be crucial. In this perspective, I will explore balances between synchrotron optical engineering choices and expansion factor, propose methods to successfully implement ExxRM in the context of human brains, and estimate how much it would cost to image the human brain in this way. Imaging brains via ExxRM may represent a crucial paradigm shift in connectomics which paves the way for holistic understanding of human brain function.
Introduction
Nanoscale connectomic imaging of the entire human brain represents a long sought-after goal that could provide the foundation for dramatic advances in neurobiology, neurotechnology, and artificial intelligence.1,2 Currently, the leading method for nanoscale connectomics is volume electron microscopy (EM). But imaging a 1 mm3 volume of mouse cortex over a period of 6 months required a tremendous collaborative effort by Yin et al. to develop a parallelized and fully automated transmission electron microscopy (TEM) system consisting of six instruments working in parallel.3 Each of these six instruments cost $125,000. The mouse brain has a volume of roughly 500 mm3, meaning that if these numbers were directly scaled, the process would take 250 years.4 That said, Yin et al.’s TEM dataset had high resolution with 4×4×40 nm voxels, so throughput might be increased by imaging at somewhat lower resolution. As such, it is conceivable to argue that advances in EM technology may enable imaging of an entire mouse brain over the course of a few years. EM therefore represents a viable option for mouse brain connectomics. But the human brain’s volume is roughly 1200 cm3, around 2400-fold larger than the mouse brain.5 Even if EM technology somehow advances to the point where an entire mouse brain can be imaged in a single year for $10M, mapping a human brain with comparable parameters would take thousands of years. In my view, this provides a strong argument for the idea that a radically different approach is necessary for dense human brain connectomics and perhaps for dense connectomics in general.
Expansion light-sheet fluorescence microscopy (ExLSFM) represents a promising alternative to EM, yet this modality also falls short when considering the volume of the human brain (particularly after expansion). For instance, Lillvis et al. utilized 8-fold expansion and lattice light-sheet microscopy (ExLLSM) to image the Drosophila central complex in three colors with approximately 15×15×50 nm (including gain from expansion) voxels over the course of 5 days.6 But even accounting for the 8-fold expansion (512-fold volume increase), this amounts to a volume of less than 0.5 mm3. Imaging an 8-fold expanded mouse brain assuming these numbers would take 7009 years. Lattice light-sheet microscopes typically cost a few hundred thousand dollars each7 and might be parallelized enough to image an entire mouse brain within a year, particularly if further hardware optimization is achievable. Yet the 2400-fold larger volume of the human brain relative to the mouse brain indicates that ExLLSM human brain connectomics may still lay out of reach. Though some investigators have made strides in combining neuronal barcoding and ExLSFM and thus might be able to image at lower resolution, allowing human brain connectomics in a reasonable time frame through parallelization of thousands of light sheet microscopes instead of tens of thousands or more, there is still room for circumventing such expensive brute-force approaches. New methods are needed to facilitate faster dense human brain connectomics.
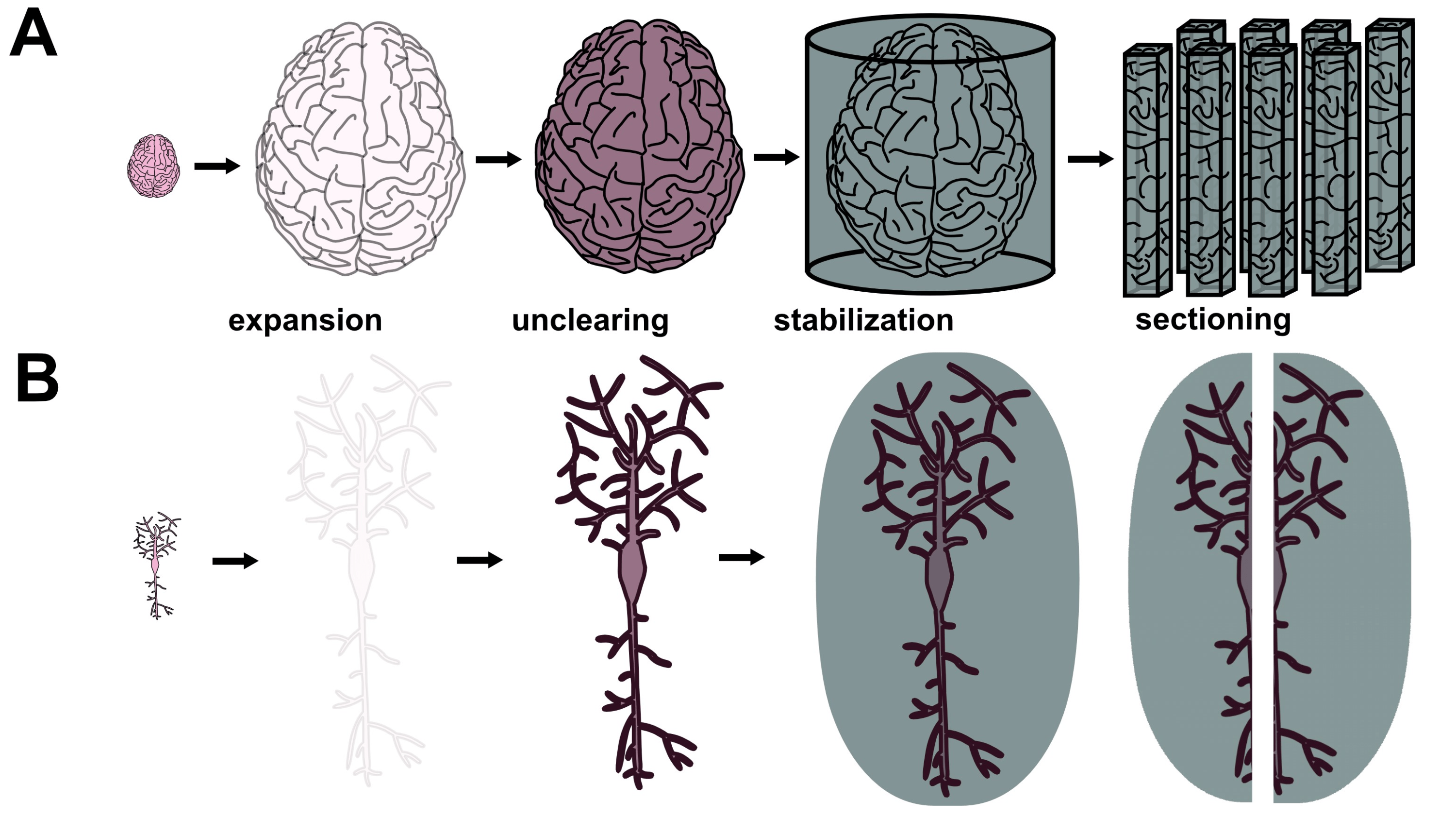
I suggest that a radically different strategy is needed to put the goal of human brain connectomics within reasonably close reach. Uniting synchrotron x-ray microtomography (XRM) (Figure 1A-B), expansion microscopy (ExM), and a recent staining method known as Unclearing Microscopy8 may facilitate ExxRM imaging of the human brain at sub-30 nm voxel size on timescales of around 1 year for a cost of around $10M. Success of this approach will necessitate overcoming some technical hurdles, yet I am optimistic that these particular challenges can be conquered. ExxRM may represent a feasible platform to acquire images suitable for dense connectomics across entire human brains.
Figure 1 Principles of synchrotron XRM. (A) Synchrotrons generate electrons from a source, propel them through a linear accelerator (linac), raise their energy in a booster ring, and then keep the electrons circulating for long periods of time at relativistic speeds in the storage ring. As relativistic electrons move along a curved path controlled by bending magnets, they emit brilliant x-ray beams in the direction tangent to the curve in the direction of the electron movement. Insertion devices that inject the x-rays into beamlines are placed at straight sections of the synchrotron ring. These insertion devices stimulate emission of bright and coherent beams into the experimental stations. (B) Microtomography beamlines receive an x-ray beam from an insertion device and filter out a narrow band of wavelengths using a monochromator. The beam passes through a sample on a rotating tomography stage that can be positionally adjusted to change the location of the field of view (FOV) within the sample. Projection images are taken across 360° of rotation. X-rays passing through the sample are converted to visible light using a scintillator, then directed by mirrors to a lens system that magnifies the image. This light hits a detector camera and data is recorded for 3D reconstruction.
Recommended methodologies for ExxRM
ExxRM’s success will require developing methodological strategies to mitigate technical challenges. Obtaining sufficient contrast to resolve subcellular features will be vital. ExM cubically dilutes the amount of cellular material per unit volume, so creative staining techniques will be needed. Approaches that ensure stability of expanded tissues under brilliant x-ray illumination for long durations will also be crucial. Multicolor imaging would greatly benefit the usefulness of whole brain connectomics data, so ways of efficiently obtaining tomograms in multiple colors are needed. These challenges must be conquered to translate ExxRM.
A central engineering hurdle for ExxRM is attaining sufficient feature contrast for capture of clear images. Because expanded tissues experience cubic dilution of target biomolecules, contrast from traditional stains such as osmium tetroxide almost entirely vanishes during x-ray microtomographic imaging (Collins et al., unpublished data). An elegant solution to this problem has come in the form of a recently developed technique called Unclearing Microscopy.8 For this technique, M’Saad et al. biotinylated primary amines (found on proteins, phosphatidylethanolamine lipids, etc.) throughout expanded samples, treated the sample with streptavidin horseradish peroxidase (streptavidin-HRP) fusion protein, and then stained with ionic silver reagents (from the EnzMet™ HRP Detection Kit) or with 3’3-diaminobenzidine (DAB). This triggered enzymatic deposition of enough chromogenic silver or DAB to make 20-fold expanded HeLa cells visible to the naked eye despite their 8000-fold increase in volume relative to the unexpanded state. Phase contrast light microscopy subsequently revealed subcellular features such as mitochondrial cristae, nuclear pore complexes, and nuclear membrane. Unclearing thus facilitates physical reconstruction of the structures that are pulled apart by ExM, filling in the gaps left by the expansion process (Figure 2A-B). Silver stain Unclearing could enable either absorption XRM or phase contrast XRM of expanded tissues since silver has excellent x-ray attenuation index β at relevant beam energies as well as excellent x-ray phase decrement δ. Focusing on phase contrast XRM may represent a better option since it can decrease the necessary dose of radiation per unit time by using x-ray wavelengths which are not absorbed as strongly by the tissue. Furthermore, phase contrast XRM is most sensitive to differences in sample density9 and silver staining forms dense precipitates of metallic silver (which in pure form has a high density of 10.49 g/cm3), so this approach might provide superior contrast in the context of ExxRM. Here, ExM’s sample dilution might prove advantageous because it could generate strongly distinct densities between silver-stained cellular features and the rest of the stabilized hydrogel. It should be noted that, for phase contrast XRM, the previously mentioned stabilization approach would need to fill the space between cellular features with a substance that differs substantially in density from the silver (or similar stain). Combining Unclearing Microscopy with phase contrast XRM could provide excellent feature contrast for ExxRM connectomics.
Expanded tissues are known for their fragility and synchrotron x-rays are known for their harshness, so strategies for solving the problem of sample degradation are needed. Newer expansion recipes which use high monomer concentrations have displayed substantially greater physical sturdiness than earlier generations of ExM hydrogels,10,11 so this may aid in stabilization to some degree. Yet additional advances in ExM sample preparation may still be necessary due to high required fluxes and long imaging times. Cutting human brain samples into smaller volumes on the order of a few centimeters may improve the situation by decreasing imaging times per sample. Keeping the sample at cryogenic temperatures with the help of cryoprotectants to prevent ice crystals damaging the tissue may also help since heat generation from the x-rays is particularly problematic for hydrated samples.12,13 That said, low temperatures would not fix the issue of radiation damage from ionization, so this issue should still be considered.14 Further stabilization might come in the form of infusion of a rapidly crosslinkable or crystalizing substance which irreversibly locks all biomolecules into place (Figure 2A-B). Polar uncharged fixatives such as glycol methacrylate may serve this purpose since they likely could substitute for water to facilitate expansion (Collins and Kohman, personal correspondence). Regardless of the specific reagents, it should be stated that the chosen substance would need to undergo an inducible crosslinking reaction that does not cause distortion of the expanded tissue. The chosen substance would need to undergo an inducible crosslinking or crystallization reaction that does not cause distortion of the expanded tissue, to not shrink or distort the expanded tissue while it diffuses into the hydrogel, and to minimally absorb x-rays at the energy range used for imaging, and to contrast sharply with the chosen stain material. So, the phase decrement of the stabilizing substance should be similar or lower compared to amorphous ice. This approach may enable sufficient stabilization to prevent nanoscale tissue degradation even with exposure to extremely brilliant synchrotron x-rays.
Figure 2 Proposed sample preparation technique for ExxRM. (A) Macroscale view of a human brain undergoing expansion, Unclearing, and stabilization. (B) Nanoscale view of a neuron within the brain undergoing expansion, Unclearing, and stabilization. Considerable amplification of signal density after Unclearing should occur.
Multicolor ExxRM would greatly enhance the value of acquired image data since synapses and key biomolecules could then be tagged within the 3D reconstruction.15,16 Fortunately, multicolor absorption x-ray microscopy represents an established technique wherein two or more staining materials undergo two or more rounds of imaging at beam energies corresponding to the absorption edges of the chosen materials. The absorption edges in question must be sufficiently distinct that minimal overlap in detection occurs across the different beam energy imaging rounds. As an example, Depannemaecker et al. employed this approach by using antibody-linked gold nanoparticles to mark neuronal nuclei in mouse brains while also staining neurons with silver via Golgi’s method.15 They imaged at a beam energy corresponding to an absorption edge of silver and at a beam energy corresponding to an absorption of gold. In addition, multicolor phase contrast x-ray imaging might be accomplished by using molecularly targeted tags with distinct densities relative to the Unclearing silver stain. For instance, gold has a density about twice that of silver, so it might be possible to segment gold nanoparticles after data acquisition. A potential challenge for these methods comes from the difficulty of inducing reliable diffusion of large metallic nanoparticles into tissue. That said, post-expansion pre-stabilization staining could help overcome the problem since expanded gels are typically more porous than pre-expansion tissue. Multicolor ExxRM in 2-3 colors should be achievable, opening doors to more useful whole-brain image datasets.
Though some trial and error will doubtless prove necessary for ExxRM optimization, the required technologies lay within fairly close reach. High contrast could be achieved by applying Unclearing Microscopy to counter signal dilution from expansion. Stability under brilliant x-rays may be possible by leveraging stabler expansion recipes with higher monomer concentrations, by cutting the brain into centimeter-scale subvolumes to decrease radiation dose per sample, by employing cryogenic temperatures during imaging, and by developing rigidity-enhancing materials to infuse into expanded gels. Multicolor ExxRM could be achieved by staining with metallic nanoparticles linked to affinity reagents and performing additional scans tuned to the absorption edges of the chosen metals. These directions may provide a path towards successful ExxRM.
How fast can synchrotrons image expanded brains?
Synchrotron facilities offer bright and coherent x-rays that can rapidly image large volumes of tissue at high resolutions (Figure 1A-B). In this regard, a recent international connectomics consortium known as SYNAPSE (Synchrotrons for Neuroscience: an Asia-Pacific Scientific Enterprise) has highlighted the value of synchrotron XRM for imaging large animal brains.17 Unfortunately, SYNAPSE does not employ ExM and is thus limited to 300 nm resolution, making their connectomics approach unsuitable for dense reconstruction of neuronal connections. At this resolution, they can only roughly reconstruct neuronal projections and certainly cannot identify synapses. Nonetheless, SYNAPSE has made technical strides towards optimizing the engineering of synchrotron beamlines towards whole-brain imaging applications. In particular, they report acquiring x-ray projection images in as little as 1 millisecond by utilizing a high-photon-energy cut-off mirror and certain filters in place of a monochromator (and “white” radiation is employed).17,18 Based on the Crowther limit,12 the number of projections Nθ needed to completely reconstruct a single tomogram is described by the equation Nθ = (π/2)N where N is the number of pixels along the horizontal direction of the image. So, for an image with N = 1000, the tomogram will require up to 3142 projection images. If each projection takes 1 ms to acquire, this means that one could potentially obtain such a tomogram in just over 3 seconds. Importantly, one should realize that larger tomograms will overall increase imaging speed despite necessitating more projections since the tomogram size scales volumetrically and the projection number only scales linearly. So, the largest possible detectors should be utilized given engineering constraints (described in next paragraph). With these metrics, one can calculate whether synchrotron microtomography might image entire expanded brains and help determine what expansion factor to use for reaching sub-30 nm voxel size while still imaging over reasonable timeframes.
Given the information described above, one may estimate optimal parameters for ExxRM (Table 1, Supplemental Code File 1). To achieve sub-30 nm pixel size, a recently developed form of ExM known as “Magnify”, which features lipid and protein retention, high physical stability, and 11-fold expansion of brain tissue may represent an ideal technique.10 At 11-fold expansion and 300 nm pixel size, effective voxel size would reach 27 nm, sufficient for dense connectomics. To image rapidly enough to acquire the entire human brain in one year, tradeoffs between detector size and pixel size must be considered. Synchrotron XRM typically uses a scintillator to convert x-rays into visible light prior to magnification via a visible light objective lens, which means the field of view (FOV) is limited by this objective lens and by how many sufficiently small pixels can be packed onto a detector. Generally, the lower the magnification of the objective lens, the wider its FOV. Yakovlev et al. demonstrated a technical accomplishment showing the feasibility of combining large FOV with high resolution.19 They developed a custom objective lens with a magnification of 6.2× and coupled it to a detector with 3.1 µm pixel size and FOV of 5 mm by 3.5 mm (10000 pixels by 7096 pixels). This enabled reconstruction of tomograms with diameter of 5 mm, height of 3.5 mm, and isotropic voxel size of 500 nm. Similar engineering of detectors and objectives might be used for mouse brain connectomics. But for human brain connectomics, one would likely need to focus on constructing a massive detector while maintaining 300 nm pixels to circumvent the probably much greater challenge of developing an objective lens with a FOV in excess of 75 mm. Imaging the entire human brain in less than one year at 27 nm effective voxel size would necessitate either a detector consisting of 62000×62000 300 nm pixels (18.6 mm × 18.6 mm and a tomogram volume of volume of about 5054 mm3)or an even vaster detector with larger pixels coupled to an objective lens with a perhaps unrealistically massive FOV. Projections acquired with such a device would require 97389 projections per tomogram according to Crowther’s limit.12 While this envisioned technology still falls beyond what has previously been achieved even in the case of Yakovlev et al., it is conceivable that current or near-future technology might allow such an objective-detector system to be commissioned for just a few million dollars, particularly given that emerging commercial smartphone cameras can now employ 200 million pixels of 560 nm and similar systems with smaller pixels are under development.20 Effective detector size might also be improved if a smaller detector panel is tiled across a larger light beam (e.g. take four images per projection of a single detector at four different tiled positions). This would linearly increase imaging time but could lead to sizable increases in overall volume acquisition speed. That said, the impact of transition time would need to be considered and might represent a significant issue. It should be noted that camera bandwidth for data recording and transmission, minimizing transition time for moving the sample between tomograms, and the time cost of slightly overlapping tomogram volumes will also need to be considered. Combining a large enough detector system with sufficiently powerful x-ray beam flux per mm2 to achieve 1 millisecond projection acquisition times (as described by the SYNAPSE consortium17), may make a human brain connectome image dataset with 27 nm effective voxel size obtainable in approximately one year of continuous imaging.
Table 1 Comparison of parameters necessary for ExxRM acquisition of a human brain connectome image dataset in 1 year at 27.27 nm effective voxel size with a mouse brain connectome image dataset in 1 month at 27.27 nm effective voxel size.
As an additionally useful metric for evaluation, Du et al. offer a mathematical model to estimate the number of photons needed for imaging a sample with sufficient contrast to distinguish between voxels of desired size.12 The model starts by considering how many photons n̄pixel are needed to image a single pixel on a tomographic slice. The pixel is assumed to contain a “feature” material with a phase decrement δf that must be distinguished against a background with a phase decrement δb. Distinctions can be made between adjacent background pixels b, overlaying background pixels bo’ and underlaying background pixels bu’. The SNR is assumed to equal 5 as based on the Rose criterion, which qualitatively represents an acceptable SNR for visually useful images. Background attenuation coefficient is given by µb’ and the thicknesses of the overlaying and underlaying background material are tb’o and tb’u respectively. After the value of n̄pixel is determined, the value is multiplied by the square N2 of the number of pixels N on the detector’s horizontal axis to find the number of photons needed per tomogram. As Du et al. explain, the number of slices is not used as a multiplicative factor since the photons are distributed across all of the rotation angles for each pixel. These calculations act as a guide to what sample properties and beam properties are needed for ExxRM.
The Du et al. model can help determine the feasibility of ExxRM by estimating the necessary flux to image the expanded brain at a desired voxel size in a reasonable time frame. Consider as an example a cube-shaped 0.3 cm3 piece of brain tissue expanded 11-fold to a block with dimensions of 3.3×3.3×3.3 cm. Based on this, assume that the average thickness of overlaying and underlaying background material is 1.65 cm (though the actual value will of course vary with the depth into the sample). Also assume that the background material is made of amorphous water ice with a density of 0.92 g/cm3. By using the Unclearing Microscopy technique, expanded features might be “filled in” after a metallic silver stain.8 Thus, I will assume that the 11-fold expanded features are made up of metallic silver with a density of 5.25 g/cm3, half the density of pure metallic silver (to account for imperfect staining). Feature phase decrements δf, background phase decrements δb, and background attenuation coefficients µb’ across x-ray energies ranging from 10 keV to 30 keV can be obtained using the calculator available at (https://henke.lbl.gov/optical_constants/pert_form.html) which is based on the Henke dataset.21 This calculator does not directly give µb’, but it does give attenuation index β, for which µb’ = 4πβ/λ. According to the resulting model (Supplemental Code File 2), the total number of photons needed to image a single tomogram (volume of 5054 mm3) using the described 18.6 mm × 18.6 mm detector within the given 11-fold expanded block at an energy of 30 keV is about 1×1014, which is within the capabilities of some contemporary synchrotrons, particularly since the flux per mm2 comes out to roughly 2.89×1011 photons/mm2 after dividing by the area of the detector.
Du et al. also give an equation (see below) for estimating the radiation dose Df imparted to a given feature in units of Gray (Gy) where 1 Gy equals 1 J of energy per kilogram of material. Leveraging the model described above (Supplemental Code File 2), this would give 0.0032 Gy as the dose imparted to a feature or 3.46×1011 Gy per 5054 mm3 tomogram. Though past studies have successfully imaged small specimens subject to comparable doses of radiation,22 this still represents a higher level of exposure than is desirable. As such, the stabilization measures (beyond just keeping the sample at cryogenic temperature during imaging) described earlier might be necessary to maintain sample integrity. Gathering experimental data on tolerable radiation dose for this specific situation will represent a vital step to clarify what engineering approaches are needed to circumvent radiation issues.
Cost estimates for whole brain ExxRM
While the power of the synchrotron facility comes with a high price tag, ExxRM may still represent the most overall cost-effective option for human brain connectomics. Consider the costs associated with the DLS as an illustrative example. The DLS is a third-generation facility and is currently one of the better synchrotrons in terms of its ability to produce bright and coherent x-rays. Building the DLS and its first seven beamlines from 2003-2007 cost about $316M, its later upgrades cost $144M and $134M, and its yearly operational costs have increased from $28M in 2007-2008 to $81M in 2019-2020 (Table 2). Based on these data points, construction of a new synchrotron beamline costs approximately $10M and yearly maintenance may cost roughly $500K. This provides a framework for estimating the cost of a dedicated human brain ExxRM connectomics beamline.
Table 2 Costs associated with the Diamond Light Source23 as a case study on how much money is needed to build and maintain a state-of-the-art synchrotron facility.
Compact Light Source (CLS) technology should be considered as well before continuing. CLS instruments produce x-ray beams that fall somewhere between laboratory x-ray microscopes and synchrotrons in terms of brightness and coherence.24 CLS instruments are furthermore small enough to fit into a single room and are inexpensive enough that a large number of them could potentially be constructed in parallel. At first glance, CLS technology seems a more economically viable alternative to synchrotron beamlines, yet it still probably is not a good option at this time. Existing CLS instruments are not likely suitable for human brain (or even mouse brain) ExxRM connectomics in the foreseeable future because their optical engineering requirements and mediocre level of x-ray flux preclude rapid tomography at submicron resolution, particularly when a large field of view is desired.25 There is a small possibility that future advances in CLS technology could change this situation, yet this seems fairly unlikely, so synchrotron-based imaging remains the best route.
Building an entire synchrotron solely for ExxRM connectomics is probably less efficient than establishing an agreement with an existing synchrotron to construct a connectomics beamline. While one might envision additional parallelization through custom design of the beamline to split the beam to land in multiple sample chambers, splitting the beam would divide the photon flux and therefore increase imaging times for no net gain in speed. As such, parallelization would likely require an additional insertion device and thus an entire new beamline for each new sample chamber. Insertion devices consist of a series of precisely engineered magnets built into straight sections of the synchrotron’s ring. These magnets, known as undulators or wigglers depending on the specifics of the insertion device, stimulate directed emission of a brilliant x-ray beam out from the storage ring.26 Yet even if we assume $10M total plus yearly maintenance costs for each connectomics beamline, imaging multiple human brains over the course of a year or a single human brain in just a few months remains a reasonable proposition. For a project as important as mapping the human brain at sufficient resolution for dense neuronal reconstruction, price tags in the range of tens of millions of dollars may not be out of reach.
Conclusion and outlook
Connectomics needs a technological paradigm shift if it is to feasibilize dense mapping of one or more human brains. Though it comes with some technical challenges, ExxRM may represent a key strategic shift that drastically reduces human brain connectomics timelines from centuries down to 1-2 years for the image acquisition step. Data storage and early processing steps will of course require data centers and high-performance computing, but this field is rapidly advancing and may indeed be capable of handling the challenge. New algorithmic advances and machine learning technologies may help tomographic reconstruction keep pace with imaging data acquisition. Assuming 1 byte per voxel, the amount of storage needed for an 11-fold expanded human brain with 300 nm physical voxel size (27 nm effective voxel size) is about 61 exabytes. Segmentation of the human brain dataset will probably represent a vastly more difficult problem as well as require substantially more compute resources, so further advances in this area will need to occur in parallel with ExxRM development. Realizing the benefits of connectomics in the form of complete computational models of the brain will take additional extensive research. Precisely correlating gene expression and electrophysiological properties with neuronal morphology (i.e. “cell type”) may represent a major step towards bridging the divide between structural data and functional activity. Nonetheless, the allure of having a holistic anatomical picture of the brain may serve as a driving force in the meantime, especially given the clinical implications of this connectome. ExxRM has the potential to transform the dense connectomics field, enabling anatomical imaging of the entire human brain with sub-30 nm voxel size in around a year for a price of roughly $10M.
Supplemental code
Copy and paste each code into a MATLAB script and run,modify as needed:
Supplemental Code 1
% optimization of ExxRM tomographic metrics calculations
% vary detectorpixels_on_a_side
detectorpixels_on_a_side = 62000; %[62000 human; 4334 mouse]; % assume that large detectors and big beams are possible
dponas = detectorpixels_on_a_side; % square detector pixels on a side, see above
pixelsize = 300; % nm; assume this size can be achieved
npixels_slice = dponas.^2; % total number of pixels on n x n detector
projections = dponas*(pi/2); % based on Du et al. equation 7, Crowther's limit
% vary pixelsize, expansionfactor
singleprojectiontime = 0.001; % seconds; very fast, as seen in https://doi.org/10.1016/j.physrep.2022.11.003 and https://doi.org/10.1107/S1600577521007633
% photons per pixel needed for absorption imaging of a feature f
square_detector_side_pixels_mm = 62000; % assume 300 nm pixel size on a 18.6 mm by 18.6 mm detector
npixels_tomo = npixel_phase*square_detector_side_pixels_mm^2; % Du et al. equation 8 gives total photons per tomogram
slices = square_detector_side_pixels_mm*(pi/2); % estimated number of slices required for tomographic reconstruction, given by N pixels on side length times pi/2
% Note: not every pixel contains a feature, but for now, we will assume that this is not the case
featuredensity = 1;
photons_tomo = featuredensity*npixels_tomo % total photons needed to image a tomogram in this situation
radiation_dose_npixelDf = npixel_phase.*(hc./lamda).*(muf./(rhof.*(tf.^2))).*exp(-mupb*tbo); % units of eV/g
C = (1.60218*10^-19)/1000% Multiplicative factor C to convert eV/g to J/kg
Df = C*radiation_dose_npixelDf % radiation dose in Gray (Gy) imparted to each feature. Multiplicative factor C converts eV/g to J/kg
Dftomo = photons_tomo.*Df
References
1. Collins, L. T. The case for emulating insect brains using anatomical “wiring diagrams” equipped with biophysical models of neuronal activity. Biol. Cybern.113, 465–474 (2019).
2. Koene, R. A. Fundamentals of whole brain emulation: state, transition, and update representations. Int. J. Mach. Conscious.04, 5–21 (2012).
3. Yin, W. et al. A petascale automated imaging pipeline for mapping neuronal circuits with high-throughput transmission electron microscopy. Nat. Commun.11, 4949 (2020).
4. Badea, A., Ali-Sharief, A. A. & Johnson, G. A. Morphometric analysis of the C57BL/6J mouse brain. Neuroimage37, 683–693 (2007).
5. Cosgrove, K. P., Mazure, C. M. & Staley, J. K. Evolving Knowledge of Sex Differences in Brain Structure, Function, and Chemistry. Biol. Psychiatry62, 847–855 (2007).
6. Lillvis, J. L. et al. Rapid reconstruction of neural circuits using tissue expansion and light sheet microscopy. Elife11, e81248 (2022).
7. Watkins, S. C. & St. Croix, C. M. Light sheet imaging comes of age. J. Cell Biol.217, 1567–1569 (2018).
8. M’Saad, O., Shribak, M. & Bewersdorf, J. Unclearing Microscopy. bioRxiv 2022.11.29.518361 (2022) doi:10.1101/2022.11.29.518361.
9. R A Lewis. Medical phase contrast x-ray imaging: current status and future prospects. Phys. Med. Biol.49, 3573 (2004).
10. Klimas, A. et al. Magnify is a universal molecular anchoring strategy for expansion microscopy. Nat. Biotechnol. (2023) doi:10.1038/s41587-022-01546-1.
11. M’Saad, O. & Bewersdorf, J. Light microscopy of proteins in their ultrastructural context. Nat. Commun.11, 3850 (2020).
12. Du, M. et al. Upscaling X-ray nanoimaging to macroscopic specimens. J. Appl. Crystallogr.54, 386–401 (2021).
13. Matsuyama, S. et al. Elemental mapping of frozen-hydrated cells with cryo-scanning X-ray fluorescence microscopy. X-Ray Spectrom.39, 260–266 (2010).
14. Lombi, E. et al. Fast X-Ray Fluorescence Microtomography of Hydrated Biological Samples. PLoS One6, e20626 (2011).
15. Depannemaecker, D. et al. Gold Nanoparticles for X-ray Microtomography of Neurons. ACS Chem. Neurosci.10, 3404–3408 (2019).
16. Logan Thrasher Collins, Kayla Siletti, Kristine Fischenich, Jennifer Coulombe, Nathan Anderson, M. S. Structural brain mapping using antibody-conjugated gold nanoparticles and x-ray microscopy. in Society for Neuroscience Conference (2019).
17. Stampfl, A. P. J. et al. SYNAPSE: An international roadmap to large brain imaging. Phys. Rep.999, 1–60 (2023).
18. Chen, H. H. et al. High-resolution fast-tomography brain-imaging beamline at the Taiwan Photon Source. J. Synchrotron Radiat.28, 1662–1668 (2021).
19. Yakovlev, M. A. et al. A wide-field micro-computed tomography detector: micron resolution at half-centimetre scale. J. Synchrotron Radiat.29, 505–514 (2022).
21. Henke, B. L., Gullikson, E. M. & Davis, J. C. X-Ray Interactions: Photoabsorption, Scattering, Transmission, and Reflection at E = 50-30,000 eV, Z = 1-92. At. Data Nucl. Data Tables54, 181–342 (1993).
22. Tennant, D. Soft X-ray microscopy with a cryo scanning transmission X-ray microscope: I. Instrumentation, imaging and spectroscopy. J. Microsc.197, 68–79 (2000).
24. Gradl, R. et al. Propagation-based Phase-Contrast X-ray Imaging at a Compact Light Source. Sci. Rep.7, 4908 (2017).
25. Gunther, B. et al. The versatile X-ray beamline of the Munich Compact Light Source: design, instrumentation and applications. J. Synchrotron Radiat.27, 1395–1414 (2020).